The Project Gutenberg eBook of A Review of the Middle American Tree Frogs of the Genus Ptychohyla
most other parts of the world at no cost and with almost no restrictions
whatsoever. You may copy it, give it away or re-use it under the terms
of the Project Gutenberg License included with this ebook or online
at www.gutenberg.org. If you are not located in the United States,
you will have to check the laws of the country where you are located
before using this eBook.
Title: A Review of the Middle American Tree Frogs of the Genus Ptychohyla
Author: William Edward Duellman
Release date: February 26, 2011 [eBook #35413]
Most recently updated: January 7, 2021
Language: English
Credits: Produced by Chris Curnow, Tom Cosmas, Joseph Cooper and
the Online Distributed Proofreading Team at
https://www.pgdp.net
*** START OF THE PROJECT GUTENBERG EBOOK A REVIEW OF THE MIDDLE AMERICAN TREE FROGS OF THE GENUS PTYCHOHYLA ***

Museum of Natural History
A Review of the Middle American Tree Frogs
of the Genus Ptychohyla
BY
WILLIAM E. DUELLMAN
University of Kansas
Lawrence
1963
University of Kansas Publications, Museum of Natural History
Editors: E. Raymond Hall, Chairman, Henry S. Fitch,
Theodore H. Eaton, Jr.
Volume 15, No. 7, pp. 297-349, pls. 11-18, 7 figs.
Published October 18, 1963
University of Kansas
Lawrence, Kansas
PRINTED BY
JEAN M. NEIBARGER, STATE PRINTER
TOPEKA, KANSAS
1963
29-6531
of the Genus Ptychohyla
WILLIAM E. DUELLMAN
| Introduction | 301 |
| Acknowledgments | 301 |
| Materials and Methods | 302 |
| Analysis of Data | 303 |
| External Morphology | 303 |
| Color and Pattern | 307 |
| Osteology | 307 |
| Tadpoles | 310 |
| Breeding Call | 312 |
| Systematic Accounts | 314 |
| Ptychohyla Taylor, 1944 | 314 |
| Key to Adults | 315 |
| Key to Tadpoles | 315 |
| Ptychohyla euthysanota Group | 315 |
| Ptychohyla euthysanota | 315 |
| Ptychohyla euthysanota euthysanota (Kellogg) | 315 |
| Ptychohyla euthysanota macrotympanum (Tanner) | 320 |
| Ptychohyla leonhardschultzei (Ahl) | 323 |
| Ptychohyla spinipollex (Schmidt) | 327 |
| Ptychohyla schmidtorum Group | 331 |
| Ptychohyla schmidtorum | 331 |
| Ptychohyla schmidtorum schmidtorum Stuart | 331 |
| Ptychohyla schmidtorum chamulae Duellman | 334 |
| Ptychohyla ignicolor Duellman | 337 |
| Distribution and Ecology | 340 |
| Geographic Distribution of the Species | 340 |
| Habitat Preference | 342 |
| Interspecific Competition | 343 |
| Reproduction and Development | 344 |
| [Pg 300]Phylogeny of Ptychohyla | 345 |
| Ptychohyla as a Natural Assemblage | 345 |
| Generic Relationships | 346 |
| Interspecific Relationships | 347 |
| Literature Cited | 349 |
Probably no ecological group of hylid frogs (some Hyla plus
Plectrohyla and Ptychohyla) in Middle America is so poorly known
as those species that live in the cloud forests on steep mountain
slopes and breed in cascading mountain streams. During the last
half of the nineteenth century most of the species of hylids living in
the lowlands of southern México and northern Central America
were named and described. Despite the extensive collecting by
Salvin and Godman, Nelson and Goldman, and the various expeditions
of the Mission Scientifique, no members of the genus Ptychohyla
were obtained until 1927, when in the mountains of El Salvador
Ruben A. Stirton found a small tree frog that subsequently was
described and named Hyla euthysanota by Kellogg (1928). Until
recently frogs of this genus were known from few specimens and in
the literature by nearly as many names.
Although I first collected Ptychohyla in 1956, it was not until 1960
that special efforts were made to obtain specimens of this genus.
The summer of 1960 was spent in southern México and Guatemala,
where every accessible stream in the cloud forests was searched for
tree frogs, especially Ptychohyla and Plectrohyla. Similar, but less
extensive, investigations were carried out in 1961 and 1962. The
result of this field work is a rather large collection of Ptychohyla
representing all of the known species, plus tape recordings of the
breeding calls and tadpoles of all of the species.
Previously, I have discussed the nomenclature of one of the
species (Duellman, 1960) and have described two new species
(Duellman, 1961). In the latter paper I made reference to a future
account (this one) that would deal with the systematics and biology
of the entire genus. Although I have series of specimens, tadpoles,
osteological preparations, and recordings of breeding calls, thereby
having a wide array of data at my disposal, much still remains to be
learned about these frogs, especially about various aspects of their
life histories. Even the validity of the genus is open to question;
this problem is discussed at length in the section beyond entitled
"Ptychohyla as a Natural Assemblage."
I am indebted to the following persons for permitting me to examine specimens
in their care: Miguel Alvarez del Toro, Museo Zoología de Tuxtla
Gutierrez, México (MZTG); Charles M. Bogert and Richard G. Zweifel, American
Museum of Natural History (AMNH); Doris M. Cochran, United States
[Pg 302]
National Museum (USNM); Norman Hartweg and Charles F. Walker, University
of Michigan Museum of Zoology (UMMZ); Robert F. Inger, Chicago
Natural History Museum (CNHM); Hobart M. Smith, University of Illinois
Museum of Natural History (UIMNH); Heinz Wermuth, Zoologisches Museum
Berlin (ZMB); and Ernest E. Williams, Museum of Comparative Zoology
(MCZ). The abbreviations following names of institutions will be used
throughout the text; the Museum of Natural History at the University of Kansas
is abbreviated KU.
Throughout my work on these frogs I have profited from discussions with
L. C. Stuart, who has made many valuable suggestions and with his characteristic
generosity has placed at my disposal his extensive collections of tadpoles
from Guatemala. For his aid I am indeed grateful. I am grateful to
Thomas E. Moore for tapes of breeding calls of two species.
My own field work was made more enjoyable and profitable through the
assistance of Dale L. Hoyt, Craig E. Nelson, Jerome B. Tulecke, and John
Wellman, all of whom spent many hours in often unsuccessful attempts to
collect specimens and record breeding calls of Ptychohyla. I am indebted to
many residents of México, Guatemala, and El Salvador for permission to work
on their land and for providing shelter, food, and guides. I am especially
grateful to Mr. and Mrs. Horatio Kelly of "Colegio Linda Vista" at Pueblo
Nuevo Solistahuacán, Chiapas, for a pleasant stay at their school; Jordi Juliá
Z. of the Comisión del Papaloapan, Ciudad Alemán, Veracruz, for arranging
for field work in northern Oaxaca in 1959; Walter Hannstein and Lothar
Menzel for the use of facilities at Finca La Paz, Guatemala, in 1960; Alan
Hempstead for the use of facilities at Finca Los Alpes, Guatemala in 1960
and 1961; and Julio Aguirre C. of the Instituto Tropical de Investigaciones
Científicas, San Salvador, El Salvador, for providing comfortable working quarters
and transportation and guides to the mountains in northern El Salvador.
Without the cheerful efforts of Jorge A. Ibarra, Director of the Museo Nacional
de Historia Natural in Guatemala, my field work would have been greatly restricted
during politically precarious times in that country. Permits to collect
in México were furnished by the late Luis Macías Arellano of the Dirección
General de Caza. Each of these individuals has my profound thanks for his
indispensable aid.
Field work on hylid frogs in Middle America has been supported by the
National Science Foundation, Grant NSF-G9827, and this is the 9th publication
on the results of study of the material from America.
During the course of this study I have examined 247 frogs that I assign to
the genus Ptychohyla, plus 40 lots of tadpoles and 12 skeletal preparations.
Furthermore, I have examined all of the type specimens. I have studied each
of the species and subspecies in the field and have examined from seven (P.
euthysanota macrotympanum) to 33 (P. spinipollex) living individuals of each
species.
Measurements given in the analysis of data and in the descriptions of the
species are those described by Duellman (1956). In the descriptions of living
colors the capitalized names are from Ridgway (1912). All interpretations of
osteological characters are based on specimens cleared in potassium hydroxide
and stained with alizarin red.
[Pg 303]
Recordings of the breeding calls were made with a Magnemite Portable
Tape-recorder; audiospectrographs were made on a vibralyzer (Kay Electric
Company) using normal pattern and wide bandwidth.
Data that are used to arrive at a systematic arrangement of the
species of Ptychohyla are analyzed and discussed below for the
values inherent in the analysis. These data are of some value also
in the recognition of species and subspecies but if employed for that
purpose the data must be used in combination with the keys and the
diagnoses of the individual species and subspecies.
Each of the external morphological characters used in the systematic
treatment of Ptychohyla, as well as the nature of the tongue,
is discussed below.
Size and Proportions.—Comparisons of size and certain proportions
are given in Table 1. Frogs of this genus are small; the largest
specimen examined is a female of P. euthysanota euthysanota having
a snout-vent length of 53.3 mm. The species comprising the
Ptychohyla schmidtorum group are smaller; the largest specimen
examined is a female of P. schmidtorum schmidtorum having a
snout-vent length of 38.0 mm. An analysis of the various measurements
and proportions shows few constant differences. Ptychohyla
ignicolor differs from all of the other species in having the head
slightly wider than long and the tympanum noticeably less than half
the size of eye. Ptychohyla spinipollex has a relatively narrow interorbital
distance, approximately equal to the width of the eyelid,
whereas in all of the other species that distance is much more than
the width of the eyelid.
Snout.—All species have a blunt snout. In P. leonhardschultzei
and P. ignicolor the snout is nearly square in lateral profile; in P.
schmidtorum the snout is slightly rounded above and below, and in
the other species it is rounded above. Ptychohyla leonhardschultzei
and P. spinipollex have a vertical fleshy rostral keel on the snout; in
these species, because of this keel, the snout in dorsal profile is
pointed. The nostrils are slightly protuberant in all species, and in
P. schmidtorum the internarial region is slightly depressed.
Table 1.—Variation in Certain Characters in the Species of Ptychohyla.
(Means Are in Parentheses Below the Ranges.)
|
Hand.—The species in the Ptychohyla euthysanota group have a
vestige of web between the first and second fingers; the other fingers
are about one-third webbed. Breeding males have a cluster of
horny nuptial spines on the thumb. The spines are largest in P.
spinipollex (Fig. 1) and vary in number from 35 to 66 (average
47.4) on each thumb. In the other species of the Ptychohyla
euthysanota group the spines are smaller and usually more numerous;
the numbers of spines on each thumb (means in parentheses)
in members of this group are: P. euthysanota euthysanota, 44-143
(83.8); P. euthysanota macrotympanum, 40-110 (63.0); P. leonhardschultzei,
[Pg 305]
24-80 (54.7). The species in the Ptychohyla schmidtorum
group have no web between the first and second fingers and only a
vestige of web between the other fingers. Furthermore, these
species lack nuptial spines in breeding males. Like the usual horny
excrescences on the thumbs of many species of frogs, the horny
spines on the thumbs of members of the Ptychohyla euthysanota
group are seasonal in development.
 Ptychohyla spinipollex and (B) Ptychohyla schmidtorum schmidtorum")
Fig. 1. Palmar views of right hands of (A) Ptychohyla spinipollex
(KU 58054) and (B) Ptychohyla schmidtorum schmidtorum (KU 58043). × 4.
Many workers have used the presence of a bifid subarticular
tubercle beneath the fourth finger as a diagnostic character of certain
species of hylids. Examination of the subarticular tubercles
in Ptychohyla reveals considerable intraspecific variation. Bifid
tubercles beneath the fourth finger are found in all species except
P. ignicolor, which is known from only two specimens. In P.
euthysanota euthysanota nearly 60 per cent and in P. schmidtorum
schmidtorum about 90 per cent of the specimens have a bifid
tubercle beneath the fourth finger on one or both hands. All
specimens of P. leonhardschultzei have either a bifid or double
tubercle beneath the fourth finger, and some have a bifid distal
tubercle beneath the third finger.
Feet.—Members of the Ptychohyla euthysanota group have a
weak tarsal fold, whereas in the species comprising the Ptychohyla
schmidtorum group the tarsal fold is absent. Webbing on the
foot extends to the discs of the third and fifth toes and to the base
of the penultimate phalanx of the fourth toe, except in P. ignicolor,
which has less webbing.
[Pg 306]
Ventrolateral Glands.—Breeding males develop thickened,
pigmented glandular areas on the sides of the body. In living
specimens of P. schmidtorum and P. ignicolor the glands are not
readily visible, but in preservative they show as distinctive orange-colored
areas. These glands are most distinct in P. euthysanota;
in many specimens of this species the glands are elevated above
the surrounding skin. The extent of the glands is variable (Fig.
2); probably this variability is due to different degrees of development
in individual frogs rather than to interspecific differences. All
Ptychohyla ignicolor and some P. schmidtorum chamulae have a
small, round glandular area on the chin; to my knowledge this does
not occur in the other species. Superficial examination of microscopic
preparations of the glands reveals no histological differences
between species. The glands occupy most of the thickened area
and have narrow ducts leading to the exterior. Detailed studies of
the histology will be reported elsewhere. Since the glands are
developed only in breeding males, it is surmised that the glands
are associated with some phase of the breeding activity.
 Ptychohyla euthysanota euthysanota, (B) Ptychohyla schmidtorum schmidtorum")
Fig. 2. Normal extent of ventrolateral glands in (A) Ptychohyla euthysanota
euthysanota (KU 58008), (B) Ptychohyla schmidtorum schmidtorum (KU
58037), and (C) Ptychohyla ignicolor (UMMZ 119603). × 21⁄4.
[Pg 307]
Tongue.—The shape of the tongue varies intraspecifically.
Usually the tongue is ovoid; in some specimens it is barely notched
posteriorly, whereas in others it is deeply notched, making the
tongue cordiform. Deeply notched cordiform tongues are found
in P. leonhardschultzei and P. schmidtorum; with the exception of
these two species, some individuals of all species have emarginate
tongues. Some individuals of all species have tongues that are
shallowly notched posteriorly.
The dorsum in living frogs of the genus Ptychohyla is primarily
yellowish or reddish brown, except in P. schmidtorum chamulae
and P. ignicolor in which it is green. Usually there are some darker
blotches or reticulations on the dorsum. The venter usually is
white; in P. ignicolor it is yellow. The venter is spotted in all members
of the Ptychohyla euthysanota group; the species, arranged
from least to most spotting ventrally, are: P. euthysanota euthysanota,
P. euthysanota macrotympanum, P. leonhardschultzei, and
P. spinipollex. The last two species also have bold dark spots on
the flanks. Ptychohyla schmidtorum lacks spots on the venter,
whereas P. ignicolor has small dark flecks ventrally.
Ptychohyla euthysanota and P. schmidtorum have white stripes
on the upper lips and on the flanks. All species have some form
of a pale stripe above the anus and usually rather distinct white or
pale stripes along the ventrolateral edges of the tarsi and forearms.
There are no bright or boldly marked flash-colors (colors
that are revealed only when the hind limbs are extended), except
in P. ignicolor, which has bright red flash-colors in the groin and
on the thighs. In life the iris varies from several different shades
of bronze color to deep red in P. schmidtorum schmidtorum.
The degree of metachrosis is moderate. Usually any change of
color in life consists only of change in the intensity of color. At
times when the over-all coloration is darkened some markings are
obscured.
The following description of the skull, hyoid, sternum, and
prepollex is based on a male specimen of P. spinipollex (KU
68632) that has been cleared and stained. The broad, flat skull
(Fig. 3) has a large frontoparietal fontanelle. The ethmoid is large
and has a flange laterally. The nasals are of moderate size and in
broad contact with the ethmoid, but are separated from one another
[Pg 308]
medially. The anterior half of the maxillary bears a thin, high
flange. The anterior process of the squamosal is short and widely
separated from the maxillary. The quadratojugal is a small spine-shaped
element projecting anteriorly from the ventral base of the
quadrate; the quadratojugal does not articulate with the maxillary.

Fig. 3. Dorsal aspect of skull of Ptychohyla spinipollex (KU
68632). Arrow indicates reduced quadratojugal. × 6.
The posteromedian part of the hyoid plate is calcified; from this
plate the long bony, posterior cornua (thyrohyales) extend posterolaterally.
The omosternum is calcified, widest anteriorly, and has a convex
anterior edge. The calcified xiphisternum is roughly bell-shaped
having short lateral processes anteriorly and a deep notch posteriorly.
The swollen thumb is supported by a dorsoventrally flattened
spine that does not extrude through the skin.
Variation.—In general, the skull varies little. Usually the quadratojugal
is present only as a short element attached to the quadrate,
but in one specimen of P. spinipollex the quadratojugal articulates
with the maxillary and forms a complete quadratojugal-maxillary
arch on each side of the skull. One specimen of P. leonhardschultzei
has a complete arch on one side and an incomplete arch
on the other.
Only P. spinipollex has lateral processes anteriorly on the xiphisternum;
in the other species the xiphisternum is deeply bell-shaped.
Ptychohyla schmidtorum and P. ignicolor have slightly longer
[Pg 309]
premaxillaries than the other species. The longer premaxillary is
reflected in the larger number of teeth on the bone—9 to 11 (average
10) in four specimens of P. schmidtorum and 10 teeth in one
P. ignicolor, as compared with 6 to 10 (average 7.9) in seven specimens
of the other species. The number of maxillary teeth in the
various species are: P. euthysanota euthysanota, 43; P. euthysanota
macrotympanum, 38; P. leonhardschultzei, 38 and 40; P. spinipollex,
34 and 40; P. schmidtorum schmidtorum, 37 and 43; P. schmidtorum
chamulae, 40 and 41; P. ignicolor, 43. The teeth on the premaxillary
and anterior part of the maxillary are long, pointed, and terminally
curved backwards. Posteriorly on the maxillary the teeth become
progressively shorter and blunter.
Variation in number of vomerine teeth is shown in Table 1.
Tadpoles of the genus Ptychohyla are adapted to live in mountain
streams. The bodies are streamlined, and the tails are long
and have low fins (Figs. 4 and 5). The mouths are large and
directed ventrally. Tadpoles of the two groups of species have
strikingly different mouthparts.

Fig. 4. Tadpoles of the Ptychohyla euthysanota group: (A) P. euthysanota
euthysanota (KU 60042), (B) P. euthysanota macrotympanum
(KU 60049), (C) P. leonhardschultzei (KU 68556), and (D) P. spinipollex
(KU 60053). ×2½.
 Ptychohyla schmidtorum schmidtorum")
Fig. 5. Tadpoles of (A) Ptychohyla schmidtorum schmidtorum (KU 60051),
(B) P. schmidtorum chamulae (KU 58199), and (C) P. ignicolor (KU 71716).
× 2½.
Lips of tadpoles of the Ptychohyla euthysanota group (Fig. 6
A-D) are folded laterally; there are 4⁄6 or sometimes 4⁄7 tooth-rows.
A lateral "wing" projects on either side of the upper beak. The
beaks have blunt, peglike serrations. Lips of tadpoles of the
Ptychohyla schmidtorum group (Fig. 6 E-G) are greatly expanded
and form a funnel-shaped disc; there are 3⁄3 short tooth-rows. There
is no lateral projection or "wing" on either side of the upper beak.
The beaks have long, pointed serrations.

Fig. 6. Mouthparts of tadpoles of Ptychohyla: (A) P. euthysanota euthysanota
(KU 60042), (B) P. euthysanota macrotympanum (KU 60049), (C)
P. leonhardschultzei (KU 68556), (D) P. spinipollex (KU 60053), (E) P.
schmidtorum schmidtorum (KU 60051), (F) P. schmidtorum chamulae (KU
58199), and (G) P. ignicolor (KU 71716). × 10.
Variation in certain structural details and in coloration is discussed
for each species and subspecies in the systematic accounts
that follow. Sizes, proportions, and numbers of tooth-rows are
tabulated in Table 2.
Table 2.—Comparison of Certain Larval Characters in the Species of
Ptychohyla. (Means Are in Parentheses Below the Ranges.)
|
Evidence on the pattern of development of tooth-rows indicates
that the inner rows develop first. A small tadpole of P.
euthysanota euthysanota has six lower rows and three fully developed
upper rows and only the beginning of the first (outer)
upper row. A small tadpole of P. euthysanota macrotympanum
has four upper rows and five lower rows. In a small tadpole of
P. leonhardschultzei the three upper and four lower tooth-rows are
well developed; the first upper and fifth lower rows are beginning
to develop, and the sixth lower row is absent. In small tadpoles
of P. spinipollex, the sixth lower row is poorly developed, and the
seventh row is absent; large individuals normally have seven lower
rows. A small tadpole of P. schmidtorum chamulae has 3⁄2 tooth-rows.
Breeding calls of all species and subspecies of Ptychohyla were
recorded in the field. Obtaining series of calls of Ptychohyla is
difficult because these frogs call mostly from vegetation along roaring
mountain streams and only by locating a calling frog some
distance from the water or along a quiet stretch of the stream can
good recordings be obtained. For example, four individuals of
[Pg 313]
P. spinipollex were recorded, but only one recording was sufficiently
free of background noise to be analyzed.
Analysis of breeding calls supports the division of the genus
Ptychohyla into two groups of species. The call of each member
of the Ptychohyla euthysanota group consists of a single long note,
whereas the call of species in the Ptychohyla schmidtorum group
consists of a series of short notes. Since no differences were found
between the calls of subspecies of any given species, the following
discussion of breeding calls pertains to species. These calls are described
briefly below and at greater length in the systematic accounts
farther on. Audiospectrographs of the breeding calls are
shown in Plate 11, and comparisons of the characteristics of the calls
are given in Table 3.
Table 3.—Comparison of the Breeding Calls of Ptychohyla
|
[A] Only an analysis of the long series of calls is given here; see text for explanation.

Audiospectrographs of the breeding calls of the five species of
(A)
(KU Tape No. 48), (C)
schmidtorum chamulae
No. 526).
P. spinipollex (Pl. 11A).—One long note is repeated at intervals of 45
seconds to four minutes and has an average dominant frequency of 4300 cycles
per second.
P. euthysanota (Pl. 11B).—One long note is repeated six to nine times at
intervals of 2.7 to 3.4 seconds and has an average dominant frequency of 3070
cycles per second.
P. leonhardschultzei (Pl. 11C).—One long note is repeated once after 10
to 13 seconds and has an average dominant frequency of 2750 cycles per
second.
P. schmidtorum (Pl. 11D).—The complete call consists of one short series
of notes alternating with two long series. Numbers of notes per series in one
individual having a typical call were 5-8-8-3-9-9. The average dominant
frequency of notes in the short and long series alike is 3400 cycles per second.
P. ignicolor (Pl. 11E).—The complete call consists of a short series of notes
alternating with a long series. In one complete recording the numbers of
notes in these series were 4-13-3-11. The notes in the short series have an
average dominant frequency of 2100 cycles per second, whereas the notes in
[Pg 314]
the long series have an average dominant frequency of 3150 cycles per second.
The four series of notes were given in one minute and 15 seconds.
The museum catalogue numbers of the specimens examined, together
with the localities from which they came, are listed at the
end of the account of each subspecies or monotypic species. The
localities that are represented by symbols on the distribution map
(Fig. 7) are in roman type; those that are not represented on the
map, because overlapping of symbols would have occurred, are in
italic type.
Ptychohyla adipoventris Taylor, 1944 [= Hyla leonhardschultzei (Ahl), 1934].
Diagnosis.—Small hylids having stream-adapted tadpoles and differing from
other hylid genera in having large ventrolateral glands in breeding males.
Composition.—Five species, two of which are made up of two subspecies,
arranged in two groups of species on the basis of morphological characters of
adults and tadpoles and on the basis of breeding calls.
Distribution.—Moderate elevations from southern Guerrero and northern
Oaxaca, México, to northern El Salvador and central Honduras.
| 1. | A weak tarsal fold; outer fingers one-third webbed; males having spiny nuptial tuberosities; color in life tan or brown with blotches or reticulations, never green; iris bronze colorP. euthysanota group—2 |
| No tarsal fold; outer fingers having only vestige of web; males lacking nuptial tuberosities; color in life green or brown; iris red or golden colorP. schmidtorum group—5 | |
| 2. | Chest, throat, and flanks usually having brown or black spots; no distinct white stripe on upper lip or on flanks; a faint white line usually present above anal opening; a rostral keel3 |
| Chest, throat, and flanks usually unspotted; distinct white line on upper lip and on flank present or not; white line above anal opening faint or well defined; no rostral keel4 | |
| 3. | Interorbital region much wider than eyelid; spots on throat and chest black; spots only occasionally present on belly; flanks marbled with black and white; nuptial spines small, as many as 80 on one thumbP. leonhardschultzei |
| Interorbital region about as wide as eyelid; spots on chest and throat brown or black; spots usually present on belly; flanks having round brown or black spots; nuptial spines moderate in size, conical, seldom more than 60 on one thumbP. spinipollex | |
| 4. | A distinct, broad, white lateral stripe usually present; usually a distinct white line above anal opening; a distinct white stripe on upper lipP. euthysanota euthysanota |
| No white lateral stripe; a faint white stripe above anal opening; no distinct white stripe on upper lipP. euthysanota macrotympanum | |
| 5. | A distinct, broad, lateral stripe; a white stripe on upper lip expanded to form a large spot below eye; hidden surfaces of thighs and webs of [Pg 315] feet not red in life; internarial region slightly depressed; diameter of tympanum greater than one-half diameter of eye6 |
| No lateral white stripe; no stripe on upper lip; in life dorsum green, hidden surfaces of thighs and webs of feet orange tan to bright red, and eye golden color; internarial region flat; diameter of tympanum less than one-half diameter of eyeP. ignicolor | |
| 6. | Webs of feet and posterior surfaces of thighs pale cream color; dorsum in life reddish brown; iris bright redP. schmidtorum schmidtorum |
| Webs of feet and posterior surfaces of thighs pale brown; dorsum in life green; iris reddish bronze colorP. schmidtorum chamulae |
| 1. | Lips greatly expanded forming a funnel-shaped mouth; tooth-rows 3⁄3P. schmidtorum group—2 |
| Lips folded laterally, not forming a funnel-shaped mouth; tooth-rows 4⁄6 or moreP. euthysanota group—4 | |
| 2. | Belly and mouth mottled; tail cream color heavily blotched with brown3 |
| Belly dark gray; tail cream color with dense brown flecking, giving brown appearanceP. schmidtorum chamulae | |
| 3. | Belly cream color with brown mottling; no large tubercle at each end of first lower tooth-rowP. schmidtorum schmidtorum |
| Belly grayish green with brown mottling; a large tubercle at each end of first lower tooth-rowP. ignicolor | |
| 4. | Tooth-rows 4⁄6; cream-colored crescent-shaped mark on posterior part of body bordered posteriorly by large brown mark5 |
| Tooth-rows usually 4⁄7 (sometimes 4⁄6); cream-colored crescent-shaped mark on posterior part of body usually indistinct, not bordered posteriorly by large brown markP. spinipollex | |
| 5. | Caudal musculature uniformly flecked with brown; lower tooth-rows 1-4 about equal in length to upper rowsP. euthysanota euthysanota |
| Caudal musculature having brown square blotches dorsally on anterior one-half of tail; lower tooth-rows 1-4 usually slightly shorter than upper rows6 | |
| 6. | Dorsal caudal blotches well defined and extending onto sides of tail; moderately large brown flecks on caudal fin; eye in life pale reddish brownP. leonhardschultzei |
| Dorsal caudal blotches faint, not extending onto sides of tail; small brown flecks on caudal fin; eye in life silvery bronzeP. euthysanota macrotympanum |
Three species in group; adults having moderate amount of webbing between
fingers, and tarsal fold; breeding males having spinous, horny, nuptial
tuberosities on pollex; mouths of tadpoles having lateral folds in lips and 4⁄6 or
4⁄7 tooth-rows; breeding call consisting of one long note.
Diagnosis.—Rostral keel absent; nuptial spines in males small; interorbital
region much wider than eyelid.
Plate 12 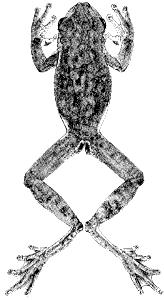 Click to View Larger. | Ptychohyla euthysanota euthysanota (Kellogg)
Hyla euthysanota Kellogg, Proc. Biol. Soc. Washington, 41:123-124, June 29, 1928 [Holotype.—USNM 73296 from Los Esemiles, Depto. Chalatenango, El Salvador; Ruben A. Stirton collector]. Mertens, Senckenbergiana, [Pg 316] 33:169-171, June 15, 1952; Abhand. Senckenbergische Naturf. Gesell., 487:29, December 1, 1952. Stuart, Proc. Biol. Soc. Washington, 67:169, August 5, 1954. Hyla rozellae Taylor, Univ. Kansas Sci. Bull., 28:78-80, pl. 9, fig. 1, May 15, 1942 [Holotype.—USNM 115039 from Salto de Agua, Chiapas, México; Hobart M. and Rozella Smith collectors]. Taylor and Smith, Proc. U. S. Natl. Mus., 95:587, June 30, 1945. Smith and Taylor, Bull. U. S. Natl. Mus., 194:86, June 17, 1948. Stuart, Proc. Biol. Soc. Washington, 67:169, August 5, 1954. Ptychohyla bogerti Taylor, Amer. Mus. Novitates, 1437:13-16, fig. 5, December 7, 1949 [Holotype.—AMNH 51847 from Río Grande, Oaxaca, México; Thomas MacDougall collector]. Stuart, Proc. Biol. Soc. Washington, 67:169, August 5, 1954. Ptychohyla euthysanota, Duellman, Univ. Kansas Publ. Mus. Nat. Hist., 13:351, April 27, 1961. |
Diagnosis.—Dorsum tan to reddish brown; venter white; rarely flecked with
brown or black; a white stripe on upper lip, on flank, and usually above anus.
Description.—The following description is based on KU 58008 from Finca
La Paz, Depto. San Marcos, Guatemala (Pl. 12). Adult male having a snout-vent
length of 35.0 mm.; tibia length, 16.5 mm.; tibia length/snout-vent length,
47.1 per cent; foot length, 14.2 mm.; head length, 11.0 mm.; head length/snout-vent
length, 31.4 per cent; head width, 10.7 mm.; head width/snout-vent length,
30.6 per cent; diameter of eye, 3.3 mm.; diameter of tympanum, 1.8 mm.;
tympanum/eye, 54.5 per cent. Snout in lateral profile nearly square, slightly
rounded above, and in dorsal profile bluntly rounded; canthus pronounced;
loreal region moderately concave; lips thick, rounded, and slightly flaring;
nostrils protuberant; internarial distance, 3.0 mm.; top of head flat; interorbital
distance, 4.1 mm., and approximately a third broader than width of eyelid,
2.9 mm. Moderately heavy dermal fold from posterior corner of eye above
tympanum to point above insertion of forelimb, covering upper edge of tympanum;
tympanum round, its diameter slightly more than its distance from eye.
Forearm moderately robust, having distinct dermal fold on wrist; dermal fold,
but no row of tubercles along ventrolateral surface of forearm; pollex only
slightly enlarged, bearing triangular shaped patch of small horn-covered spines
(128 on right, 134 on left); second and fourth fingers equal in length; subarticular
tubercles round, distal one on fourth finger bifid; discs moderate in
size, that of third finger equal to diameter of tympanum; no web between first
and second fingers; other fingers one-third webbed. Heels broadly overlap
when hind limbs adpressed; tibiotarsal articulation reaches to middle of eye;
low rounded tarsal fold; inner metatarsal tubercle large, elliptical, and flat;
outer metatarsal tubercle small and round; low dermal fold from heel to disc
of fifth toe; subarticular tubercles round; length of digits from shortest to longest
1-2-5-3-4; third and fifth toes webbed to base of disc; fourth toe webbed to
proximal end of penultimate phalanx; thin dermal fold from inner metatarsal
tubercle to disc of first toe; disc smaller than on fingers. Anal opening at the
level of the upper edge of thighs; anal flap short; anal opening bordered above
by thin transverse dermal fold and laterally by heavy dermal fold. Skin of
dorsum and ventral surfaces of forelimbs and shanks smooth; that of throat,
belly, and ventral surfaces of thighs granular. Ventrolateral glands moderately
developed, not reaching axilla or groin and broadly separated midventrally.
Tongue ovoid, emarginate, and only slightly free posteriorly; vomerine teeth
2-2, situated on small triangular elevations between ovoid inner nares; openings
[Pg 317]
to vocal sac large, one situated along inner posterior edge of each mandibular
ramus.
Dorsal ground-color of head, body, and limbs dull reddish brown with
irregular dark brown reticulations on head and body and dark brown transverse
bands on limbs; dorsal surfaces of first and second fingers and webbing on hand
cream color; dorsal surfaces of third and fourth fingers dull brown; anterior
surfaces of thighs dull creamy yellow; posterior surfaces of thighs dull brown;
tarsi and toes tan with brown flecks; webbing of feet brown; faint creamy white
stripe along lateral edges of tarsi and forearms; thin white line along edge of
upper lip; distinct white stripe above and beside anal opening; axilla white;
throat, chest, belly, and ventral surfaces of forelimbs creamy white; flanks
white, separated from pale venter by a row of partly connected dark brown
spots; ventral surfaces of thighs dull creamy yellow; feet grayish brown;
ventrolateral glands pale grayish brown; small brown flecks on periphery of
chin.
In life the dorsal ground-color was pale reddish brown (Orange-Cinnamon);
dorsal reticulations dark brown (Chocolate); dorsal surfaces of first
and second fingers and webbing on hands creamy tan (Light Pinkish Cinnamon);
posterior surfaces of thighs reddish brown (Vinaceous-Tawny); webbing
of feet gray (Deep Mouse Gray); throat and belly grayish white (Pale
Gull Gray); ventral surfaces of hind limbs creamy white (Marguerite Yellow);
spots on flanks dark brown (Warm Sepia); iris reddish bronze (Apricot
Orange).
Variation.—No geographic variation in structural characters is discernible;
variation in size and proportions is given in Table 1. Of 32 adults examined,
seven have the tongue shallowly notched posteriorly; in the others the tongue
is emarginate. Twenty specimens have a bifid subarticular tubercle beneath
the fourth finger; in the others there are no bifid tubercles.
The coloration described above is typical of the 16 specimens available from
Finca La Paz. The living coloration at night, when the frogs were collected,
was somewhat darker than the living colors described above, which were recorded
for the frogs the morning after collection, at which time one individual
had a pale reddish brown dorsum (Orange-Cinnamon) with dull olive green
(Deep Grape Green) reticulations on the back and transverse bands on the
limbs; the dorsal surfaces of the first and second fingers and the discs on the
third and fourth fingers were orange (Mikado Orange).
More than half of the specimens from Finca La Paz agree in all essential
characters with the description given above. The distinctness of the white
stripe on the upper lip is variable; in two individuals the stripe is barely discernible.
Likewise, in some individuals the white stripe on the flanks is not
distinct, either because there are few or no brown spots separating the stripe
from the pale venter, or because the ventrolateral gland has diffused the pale
color on the flanks. There is some noticeable variation in dorsal coloration,
either through a greater or lesser development of dark pigment. One specimen
(KU 58007) is grayish tan above with dark brown markings; the
posterior surfaces of the thighs are dull grayish yellow; the first and second
fingers and the webbing on the hands are pale yellowish gray; the belly and
throat are dusty white; the flecks on the throat are gray; the ventral surfaces
of the feet are grayish brown. Dark individuals, such as KU 58009 have a
uniform dark brownish black dorsum; the belly is cream; the first and second
[Pg 318]
fingers and the webbing on the hands are dull creamy tan; the dorsal and
ventral surfaces of the feet are dark brown. In KU 58013 there is a heavy
suffusion of brown on the throat and flanks. Two specimens have scattered
white flecks on the dorsum.
The reddish brown dorsal ground-color with dark brown reticulations on
the head and body and dark brown transverse bands on the limbs seems to be
rather constant throughout the range of the subspecies. Likewise, the presence
of the white stripe on the upper lip and the white stripe around the anal opening
are present on most specimens. In breeding males having well-developed
ventrolateral glands the lateral white stripe often is obliterated.
Description of Tadpole.—The following description is based on KU 60042
from Finca La Paz, Depto. San Marcos, Guatemala (Figs. 4A and 6A). No
limb buds; total length, 35.8 mm.; body length, 11.2 mm.; body length/total
length, 31.3 per cent. Body moderately depressed, slightly wider than deep,
ovoid in dorsal profile; mouth directed ventrally; eyes small, directed dorsolaterally;
nostrils slightly protuberant and directed anteriorly, closer to eye than
snout; spiracle sinistral and posteroventrad to eye; anal tube dextral. Caudal
fin low, rounded posteriorly; depth of caudal musculature about one-half
greatest depth of caudal fin; musculature extends nearly to tip of tail.
Mouth large; lips having deep lateral folds; two complete rows of papillae
on lips; five to six rows of papillae laterally. Beaks moderately developed,
bearing peglike serrations; slender lateral projections on upper beak; tooth-rows
4⁄6; upper rows subequal in length, second longest; fourth row interrupted
medially; lower rows complete; lower rows 1-4 equal in length to upper rows;
fifth lower row somewhat shorter; sixth lower row short.
Body brown above; tip of snout cream color; grayish cream color below;
caudal musculature creamy tan; caudal fin transparent; cream-colored crescent-shaped
mark on posterior edge of body and anterior part of caudal musculature,
bordered posteriorly by dark brown blotch; scattered brown flecks on caudal
musculature and posterior part of caudal fin. Eye bronze color in life.
Variation.—The variation in size and proportions is given in Table 2. In
some specimens the first upper tooth-row is irregular, sometimes broken, and
often shorter than other upper tooth-rows. Usually the fourth upper and first
lower, and sometimes the sixth lower, tooth-rows are interrupted medially. One
specimen has a short, irregular, seventh lower tooth-row; all others have six.
The cream-colored crescent-shaped mark usually is distinct. The brown
blotch posterior to this mark is variously shaped ranging from a narrow vertical
bar to a triangular blotch. Brown flecks seldom are present on the anterior
part of the ventral caudal fin.
Comparisons.—Aside from the characters given in the diagnosis, P. euthysanota
euthysanota can be distinguished from both P. spinipollex and P. leonhardschultzei
by the absence of bold black and white marbling on the flanks;
furthermore, from the former it can be distinguished by having more and
smaller horny nuptial spines and from the latter by having the snout, in
lateral profile, rounded above and not acutely angulate. Ptychohyla euthysanota
euthysanota differs from P. euthysanota macrotympanum by normally
having a darker dorsal color, broader stripe on upper lip, and a distinct lateral
stripe.
Occurring sympatrically with Ptychohyla euthysanota euthysanota are several
species of Plectrohyla, all of which differ in having a bony prepollex, rather
[Pg 319]
rugose skin on the dorsum, and more squat bodies. Other sympatric species
are Ptychohyla schmidtorum schmidtorum, which lacks a tarsal fold and nuptial
spines and has a red eye in life, Hyla salvadorensis, which has a green dorsum
and lacks spinous nuptial tuberosities, and Hyla sumichrasti, a small yellow
frog usually lacking vomerine teeth.
Life History.—This subspecies breeds in clear, swift mountain
streams. Males call from stems and leaves of plants at the edge of,
or overhanging, the streams. The breeding call consists of a soft
"wraack," repeated at intervals of three to four seconds. Each note
has a duration of 0.60 to 0.65 seconds and has 91 to 102 pulses per
second; the dominant frequency falls between 3000 and 3200 cycles
per second.
Tadpoles in various stages of development were found at Finca
La Paz, Guatemala, in late July. This indicates that there is either
extreme differential growth, or, more probably, an extended breeding
season. A tadpole having a body length of 6.8 mm. and a total
length of 19.1 mm. has a short median first upper tooth-row; lower
tooth-rows 3-6 are only two-thirds as long as lower rows 1 and 2.
Two recently metamorphosed young have snout-vent lengths of 14.2
and 14.8 mm.; they are colored like the adults.
Remarks.—The type specimen of Hyla euthysanota Kellogg
(1928:123) is a female; therefore, when Taylor (1944) proposed the
name Ptychohyla for hylids having ventrolateral glands in breeding
males, he was unaware that Hyla euthysanota was a member of this
group. In his description of Hyla rozellae, Taylor (1942) did not
compare his specimens with Hyla euthysanota, but instead placed
H. rozellae with H. loquax and H. rickardsi. The type series of H.
rozellae consists of one large adult female and several metamorphosing
young. Taylor (1949:16) based the description of Ptychohyla
bogerti on two males and compared these specimens with P.
adipoventris Taylor [= P. leonhardschultzei (Ahl)]. Thus, in a
period of 22 years the females of this species were given two names
and the male another. Stuart (1954:169) suggested that Hyla
euthysanota and Hyla rozellae were Ptychohyla. Now that sufficient
specimens are available from throughout the range it is possible to
determine that the various named populations are conspecific.
Distribution.—This subspecies inhabits cloud forests at elevations of 660 to
2200 meters on the Pacific slopes of the Sierra Madre from extreme eastern
Oaxaca and western Chiapas, México, through Guatemala to northern El
Salvador; probably it occurs also in southwestern Honduras. Aside from the
specimens listed below, three in the Frankfurt Museum from Depto. Santa Ana,
El Salvador (44571, Hacienda San José; 43040, Hacienda Los Planes; 65119,
Miramundo) are listed by Mertens (1952:29).
[Pg 320]
Specimens examined.—Mexico: Chiapas: Cascarada, 30 km. W of Cíltapec,
UMMZ 87851-2; Cerro Ovando, UMMZ 87853-4; Chicomuselo, UMMZ 94439-40;
Finca Juárez, 28 km. N of Escuintla, USNM 115052-5; Las Nubes, Cerro
Ovando, USNM 115030-8; Salto de Agua, USNM 115039-51. Oaxaca: Cerro
Pecho Blanco, UIMNH 40963; between La Gloria and Cerro Azul, UIMNH
40976-7; Río Grande, AMNH 51847-8; Santo Tomás Tecpan, UIMNH 41071.
Guatemala: San Marcos: Finca La Paz, 2 km. W of La Reforma, KU
58001-14, 59937 (skeleton), 60042-3 (tadpoles), 60044 (4 young), MCZ
34997, UMMZ 107739, 123151-7 (tadpoles); Finca Pirineos, Río Samalá,
CNHM 35066. Santa Rosa: Finca La Gloria, UMMZ 123148 (tadpoles),
123150 (tadpoles). Sololá: Finca Santo Tomás, UMMZ 123149 (tadpoles);
Olas de Mocá, near Mocá, CNHM 20208.
El Salvador: Chalatenango: Los Esemiles, USNM 73296. Santa Ana:
Miramundo, CNHM 65120.
Plate 13  Click to View Larger. | Ptychohyla euthysanota macrotympanum (Tanner)
Hyla macrotympanum Tanner, Great Basin Nat., 17:52-53, July 31, 1957 [Holotype.—AMNH 62141 (formerly BYU 13752) from 10 miles east of Chiapa de Corzo, Chiapas, México; Robert Bohlman collector]. Ptychohyla macrotympanum, Duellman, Univ. Kansas Publ. Mus. Nat. Hist., 13:351, April 27, 1961. |
Diagnosis.—Dorsum usually pale tan; venter white with scattered brown or
black flecks; a thin white stripe on upper lip and another above anal opening;
no distinct white stripe on flanks.
Description.—The following description is based on KU 58049 from Linda
Vista, Chiapas, México (Pl. 13). Adult male having a snout-vent length of
38.0 mm.; tibia length, 19.5 mm.; tibia length/snout-vent length, 51.3 per cent;
foot length, 15.7 mm.; head length, 11.8 mm.; head length/snout-vent length,
31.1 per cent; head width, 11.7 mm.; head width/snout-vent length, 30.8 per
cent; diameter of eye, 3.8 mm.; diameter of tympanum, 2.1 mm.; tympanum/eye,
55.2 per cent. Snout in lateral profile nearly square, slightly
rounded above, and in dorsal profile bluntly rounded; canthus pronounced;
loreal region concave; lips thick, rounded, and slightly flaring; nostrils protuberant;
internarial distance, 3.1 mm.; top of head flat; interorbital distance,
3.8 mm., and approximately a fourth broader than width of eyelid, 3.1 mm.
A moderately heavy dermal fold from posterior corner of eye above tympanum
to point above insertion of forelimb, covering upper edge of tympanum; tympanum
round, its diameter equal to its distance from eye. Forearm moderately
robust, having a distinct dermal fold on wrist; dermal fold, but no
row of tubercles along ventrolateral surface of forearm; pollex only slightly
enlarged, bearing triangular patch of small horn covered spines (94 on right,
100 on left); fourth finger slightly longer than second; subarticular tubercles
round, none bifid; discs moderate in size, that of third finger equal to diameter
of tympanum; vestige of web between first and second fingers; other fingers
one-third webbed. Heels broadly overlap when hind limbs adpressed; tibiotarsal
articulation reaches to middle of eye; weak tarsal fold on distal two-thirds
of tarsus; inner metatarsal tubercle large, elliptical, and flat; outer
metatarsal tubercle small and round; no dermal fold from heel to disc of fifth
toe; subarticular tubercles round; length of digits from shortest to longest
1-2-5-3-4; third toe webbed to base of disc; fifth toe webbed to middle of
penultimate phalanx; fourth toe webbed to proximal end of penultimate
phalanx; no fold of skin from inner metatarsal tubercle to base of disc on first
[Pg 321]
toe; discs much smaller than on fingers. Anal opening near upper edge of
thighs; short anal flap bordered above by thin dermal fold; small tubercles
and heavy dermal fold lateral to anal opening. Skin of dorsum and ventral
surfaces of fore limbs and shanks smooth; that of throat, belly, and ventral
surfaces of thighs granular. Ventrolateral glands weakly developed, not
reaching axilla or groin and broadly separated midventrally. Tongue ovoid,
shallowly notched posteriorly, and barely free behind; vomerine teeth 2-2,
situated on small triangular elevations between ovoid inner nares; openings to
vocal sac large, one situated along inner posterior edge of each mandibular
ramus.
Dorsal ground-color of head, body, and limbs pale pinkish tan with the
greatest part of head and body covered by large gray interconnected blotches;
black flecks over most of dorsum; grayish brown transverse bands on shanks;
dorsal surfaces of first and second fingers pale grayish yellow; dorsal surfaces
of third and fourth fingers and webbing on hand pale grayish tan; anterior
surfaces of thighs pale flesh-color; posterior surfaces of thighs pale grayish
yellow; dorsal surfaces of tarsi and toes pale grayish tan with black flecks;
webbing of feet pale gray; faint creamy white stripes along ventrolateral
edges of tarsi and forearms; a very thin white line along edge of upper lip;
a narrow grayish white stripe above anal opening; axilla gray; throat, chest,
belly, and ventral surfaces of forelimbs pale grayish white; ventral surfaces of
hind limbs cream color; flanks gray flecked with brown; ventral surfaces of
feet grayish tan; ventrolateral glands pinkish tan; anterior one-half of chin
flecked with brown.
In life the dorsum was pale tan (Pinkish Buff); the dark markings on dorsum
dull brown (Tawny-Olive); tarsi pale tan (Pale Pinkish Buff); flanks
pinkish tan (Pale Cinnamon-Pink); iris coppery bronze (Capucine Orange).
Variation.—The few specimens from a limited geographic region preclude
any analysis of geographic variation. All specimens, except the one described
above, have the fifth toe webbed to the base of the disc. Many individuals
have a bifid subarticular tubercle beneath the fourth finger. The shape of
the posterior edge of the tongue varies from nearly straight and shallowly
notched to bluntly rounded and emarginate. Two females (KU 58050-1)
have more pointed snouts in dorsal profile than do males.
Some specimens, such as KU 58048, are notably darker than the specimen
described above; in dark specimens the dorsum is brown with dark brown
markings; all fingers and the webbing on the hand are brown. The tarsi and
feet are like those in the specimen described above, but the posterior surfaces
of the thighs are yellowish tan heavily suffused with brown; the venter is
cream color. In life KU 58048 had a brown (Verona Brown) dorsum with
dark brown (Chocolate) markings. KU 58047 is slightly darker than KU
58048 and has scattered small white flecks on the dorsum. All specimens
have the thin white line on the upper lip, but in some individuals it is indistinct.
The grayish white line above the anus is present in all specimens.
Description of Tadpole.—The following description is based on KU 60049
from Río Hondo, 9.5 kilometers south of Pueblo Nuevo Solistahuacán, Chiapas,
México (Figs. 4B and 6B). No limb buds; total length, 36.2 mm.; body
length, 11.1 mm.; body length/total length, 30.6 per cent. Body moderately
depressed, slightly wider than deep, ovoid in dorsal profile; mouth directed
ventrally; eyes small, directed dorsolaterally; nostrils slightly protuberant and
[Pg 322]
directed anteriorly, somewhat closer to eye than snout; spiracle sinistral and
posteroventrad to eye; anal tube dextral. Caudal fin low, acutely rounded
posteriorly; depth of caudal musculature slightly more than one-half greatest
depth of caudal fin; caudal musculature extending nearly to tip of tail.
Mouth large; lips having deep lateral folds; two complete rows of papillae
on lips; five or six rows of papillae laterally. Beaks moderately developed,
bearing small peglike serrations; moderately wide lateral projections on upper
beak; tooth-rows 4⁄6; upper rows subequal in length; fourth row interrupted
medially; length of lower rows 1-4 subequal to upper rows; fifth and sixth
lower rows decreasing in length; first lower row interrupted medially.
Body brown above; tip of snout cream color; venter creamy white; caudal
musculature creamy tan; caudal fin transparent; cream-colored crescent-shaped
mark on posterior edge of body and anterior part of caudal musculature,
bordered posterodorsally by dark brown blotch; four dark brown blotches on
dorsum of anterior part of caudal musculature; scattered brown flecks on
caudal musculature and fin; eye silvery bronze in life.
Variation.—The variation in size and proportions is given in Table 2. All
specimens have 4⁄6 tooth-rows; in some the first lower row is interrupted
medially. Specimens from Jacaltenango and two kilometers west of San
Pedro Necta, Depto. Huehuetenango, Guatemala, have weakly developed sixth
lower tooth-rows.
The cream-colored crescent-shaped mark is distinct in all specimens; the
dorsal blotches on the anterior part of the caudal musculature are narrow and
do not extend far onto the sides of the tail. The blotches are most distinct in
small tadpoles and sometimes indistinct in large ones.
Comparisons.—Ptychohyla euthysanota macrotympanum can be distinguished
from both P. spinipollex and P. leonhardschultzei by the absence of bold black
and white marbling on the flanks, as well as by the characters given in the
diagnosis; furthermore, from the former it differs in having more and smaller
horny nuptial tuberosities and from the latter by having the snout, in lateral
profile, rounded above instead of angulate. Ptychohyla euthysanota macrotympanum
differs from P. e. euthysanota by normally having a paler dorsum, narrower
stripe on upper lip, and no distinct lateral stripe.
Ptychohyla euthysanota macrotympanum occurs sympatrically with Plectrohyla
guatemalensis and P. matudai matudai. Each of the last two has a bony
prepollex, rather rugose skin on the dorsum, and more squat body. Other
sympatric species are Hyla walkeri, which has a green dorsum with brown
markings and a rather pointed snout, and Hyla sumichrasti, a small yellow
frog usually lacking vomerine teeth.
Life History.—This species breeds in clear mountain streams in
mixed pine and broad-leafed forest. Males call from trees and
bushes along the streams. The breeding call consists of a soft
"wraack," repeated three to nine times with intervals of 2.7 to 3.4
seconds between notes. Each note has a duration of 0.60 to 0.65
second, and a rate of 92 to 100 pulses per second; the dominant
frequency falls between 3000 and 3200 cycles per second (Pl. 11B).
The call is indistinguishable from that of P. e. euthysanota.
[Pg 323]
Tadpoles in various stages of development were found in the Río
Hondo, Chiapas, on June 16, 1960. The smallest tadpole has a total
length of 24.1 mm.; in this individual the sixth lower tooth row has
barely started to develop. A metamorphosing frog taken at the
same time has a snout-vent length of 19.8 mm., a short remnant of
the tail, and the mouth and tongue developed, whereas another
individual having a snout-vent length of 17.8 mm. and a tail 31.0 mm.
in length still has larval teeth. Three completely metamorphosed
juveniles collected by L. C. Stuart at Jacaltenango, Guatemala, on
June 6 and 7, 1955, have snout-vent lengths of 16.0, 16.0, and
16.1 mm.
Remarks.—Tanner (1957:52) based the description of Hyla
macrotympanum on a single female, which, of course, lacked the
characters diagnostic of Ptychohyla. On the basis of general external
characters Tanner suggested that Hyla macrotympanum was
related to H. miotympanum, from which it differed in having a
larger tympanum and a bifid subarticular tubercle beneath the
fourth finger. The collection of additional females, together with
males of the species, has shown that Hyla macrotympanum is a
Ptychohyla.
Intergradation between Ptychohyla euthysanota and P. macrotympanum
has not been demonstrated. Currently their ranges are
separated by the dry valleys of the Río Grijalva and Río Cuilco.
The similarity in structure of the adults and tadpoles and the indistinguishable
breeding calls are the basis for considering the two
populations subspecies.
Distribution.—This species occurs in mixed pine and broad-leafed forests
at elevations of 700 to 1700 meters on the southern slopes of the Chiapan Highlands
and Sierra de Cuchumatanes, in Guatemala. These forests are on the
south facing slopes north of the valleys of the Río Grijalva and Río Cuilco.
Specimens examined.—Mexico: Chiapas: 6 km. NE of Chiapa de Corzo,
TCWC 16183; 16 km. E of Chiapa de Corzo, AMNH 62141; Linda Vista, 2 km.
NW of Pueblo Nuevo Solistahuacán, KU 58049-51, 59939 (skeleton); Río
Hondo, 9.5 km. S of Pueblo Nuevo Solistahuacán, KU 58047-8, 60046-7,
60048-9 (tadpoles); San Fernando, MZTG 15, 17; Tonina (ruins), KU 41592.
Guatemala: Huehuetenango: Jacaltenango, UMMZ 123139 (tadpoles);
0.5 km. E of Jacaltenango, UMMZ 123142-3; 2 km. S of Jacaltenango, UMMZ
123141; 2 km. W of San Pedro Necta, UMMZ 123140 (tadpoles).
Plate 14 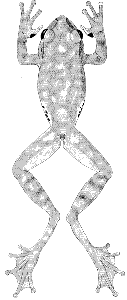 Click to View Larger. | Ptychohyla leonhardschultzei (Ahl)
Hyla leonhard-schultzei Ahl, Zool. Anz., 106:185-186, fig. 1, April 15, 1934 [Holotype.—ZMB 34353 from Malinaltepec, Guerrero, México; Leonhard Schultze collector]. Smith and Taylor, Bull. U. S. Natl. Mus., 184:87, June 17, 1948. Hyla godmani, Ahl, Zool. Anz., 106:186, April 15, 1934. Hyla pinorum Taylor, Proc. Biol. Soc. Washington, 50:46-48, pl. 2, fig. 2, [Pg 324] April 21, 1937 [Holotype.—UIMNH 25049 from Agua del Obispo, Guerrero, México; Edward H. Taylor collector]. Smith and Taylor, Bull. U. S. Natl. Mus., 194:87, June 17, 1948. Ptychohyla adipoventris Taylor, Univ. Kansas Sci. Bull., 30:41-45, May 15, 1944 [Holotype.—UIMNH 25047 from Agua del Obispo, Guerrero, México; Edward T. Taylor collector]. Smith and Taylor, Bull. U. S. Natl. Mus., 194:91, June 17, 1948. Taylor, Amer. Mus. Novitates, 1437:16, December 7, 1949. Stuart, Proc. Biol. Soc. Washington, 67:169, August 5, 1954. Hyla milleri Shannon, Proc. U. S. Natl. Mus., 101:473-477, figs. 92b, 93a-c, May 17, 1951 [Holotype.—USNM 123700 from San Lucas Camotlán, Oaxaca, México; Walter S. Miller collector]. Ptychohyla leonhard-schultzei, Duellman, Herpetologica, 16:191-197, figs. 1-3, September 23, 1960; Univ. Kansas Publ. Mus. Nat. Hist., 13:351, April 27, 1961. |
Diagnosis.—Rostral keel present; snout in lateral profile not rounded above;
interorbital region much broader than eyelid; distal subarticular tubercle
beneath fourth finger bifid or double; no white stripe on edge of upper lip;
flanks white with black spots.
Description.—The following description is based on KU 64117 from Vista
Hermosa, Oaxaca, México (Pl. 14). Adult male having a snout-vent length
of 35.6 mm.; tibia length, 18.0 mm.; tibia length/snout-vent length, 50.5 per
cent; foot length, 14.3 mm.; head length, 10.7 mm.; head length/snout-vent
length, 30.1 per cent; head width, 10.6 mm.; head width/snout-vent length,
29.8 per cent; diameter of eye, 3.6 mm.; diameter of tympanum, 1.8 mm.;
tympanum/eye, 50.0 per cent. Snout in lateral profile square, not rounded
above, and in dorsal profile rounded with pointed tip resulting from vertical
rostral keel; canthus pronounced; loreal region barely concave; lips thick,
rounded, and barely flaring; nostrils protuberant; internarial distance, 3.2 mm.;
top of head flat; interorbital distance, 3.8 mm., and approximately a fifth
broader than width of eyelid, 3.2 mm. A moderately heavy dermal fold from
posterior corner of eye above tympanum and curving ventrad to anterior edge
of insertion of forelimb, covering upper edge of tympanum; tympanum round,
its diameter equal to its distance from eye. Forearm moderately robust, having
distinct dermal fold on wrist; row of small, low, rounded tubercles along
ventrolateral surface of forearm; pollex only slightly enlarged, bearing triangular
patch of small horn-covered spines (56 on right, 62 on left); second finger
noticeably shorter than fourth; subarticular tubercles round, distal ones on
third and fourth toes bifid; discs moderate in size, that of third finger slightly
larger than diameter of tympanum; no web between first and second fingers;
other fingers one-third webbed. Heels broadly overlap when hind limbs adpressed;
tibiotarsal articulation reaches to middle of eye; a low rounded tarsal
fold on distal half of tarsus; inner metatarsal tubercle elevated, flat, and elliptical;
outer metatarsal tubercle at base of fourth toe, round; row of low, sometimes
indistinct, tubercles from heel to base of fifth toe; subarticular tubercles
round; length of digits from shortest to longest 1-2-3-5-4, third and fifth being
about equal in length; third and fifth toes webbed to base of disc; fourth
toe webbed to base of penultimate phalanx; discs of toes much smaller than on
fingers. Anal opening near dorsal surface of thighs; short anal flap; opening
bordered laterally by heavy dermal fold and ventrolaterally by large tubercles.
Skin of dorsum and ventral surfaces of forelimbs and shanks smooth; that of
throat, belly, and ventral surfaces of thighs granular. Ventrolateral glands
[Pg 325]
moderately developed, reaching axilla but not to groin and broadly separated
midventrally. Tongue cordiform, shallowly notched behind and barely free
posteriorly; vomerine teeth 4-3, situated on transverse elevations between ovoid
inner nares; openings to vocal sac large, one situated along inner posterior edge
of each mandibular ramus.
Dorsal ground-color of head, body, and limbs pale tan with large interconnected
dark brown blotches on head and body and broad dark brown
transverse bands on limbs; dorsal surfaces of first and second fingers and of
webbing of hands pale brown; dorsal surfaces of third and fourth fingers dark
brown; anterior surfaces of thighs flesh-color; posterior surfaces of thighs
brown; dorsal surfaces of tarsi and feet dark brown; narrow white stripe along
ventrolateral edges of forearms and tarsi; a faint creamy white stripe above
anal opening; axilla white; flanks creamy white, boldly spotted with black;
throat and chest white; ventral surfaces of tarsi and feet dark brown; other
ventral surfaces dusty cream color; large brown spots on chin and anterior
part of abdomen.
In life the dorsum was reddish brown (Orange-Cinnamon) with dark
brown (Chocolate) blotches; first and second fingers and webbing on hand
pale reddish brown (Cinnamon); webbing on feet dark brown (Clove Brown);
flanks pale creamy white (Pale Olive Buff) with dark brown (Bone Brown)
spots; iris reddish bronze (Apricot Orange).
Variation.—No noticeable geographic variation is apparent in the few available
specimens; variations in proportions are given in Table 1. The distal
subarticular tubercle of the fourth finger is either bifid or double in all specimens;
that on the third finger usually is bifid, sometimes single. The vertical
rostral keel is prominent in all freshly preserved specimens; in some older
specimens it is indistinct. The tongue always is notched posteriorly; in some
individuals the notch is shallow; in others it is deep.
Some specimens are paler and less boldly marked than the specimen described
above. All specimens from Agua del Obispo and some specimens
from the northern slopes of the Sierra Madre Oriental in Oaxaca have a pale
tan dorsum with brown markings. In most individuals the white color in the
axilla extends onto the posterior edge of the upper arm. The creamy white
color of the flanks is constant and usually extends slightly dorsad in the
inguinal region. The white stripe above, and sometimes continuing down beside,
the anal opening varies from a thin indistinct line or row of flecks to a
distinct continuous stripe. Two specimens have dark brown spots on the
belly; in the others the spots are confined to the flanks and throat.
Description of tadpole.—The following description is based on KU 68556
from 7.5 kilometers south of Yetla, Oaxaca, México (Figs. 4C and 6C). No
limb buds; total length, 37.3 mm.; body length, 12.2 mm.; body length/total
length, 32.7 per cent; body slightly depressed, barely wider than deep, ovoid
in dorsal profile; mouth directed ventrally; eyes small, directed dorsolaterally;
nostrils barely protuberant and directed anterolaterally, about midway between
snout and eye; spiracle sinistral and posteroventrad to eye; anal tube dextral.
Caudal fin low, bluntly rounded posteriorly; greatest depth of caudal musculature
about one-half depth of caudal fin; musculature extends nearly to tip of
tail.
Mouth large; lips having deep lateral folds; two complete rows of papillae
on lips; five to seven rows of papillae laterally; beaks moderately developed,
[Pg 326]
bearing short peglike serrations; moderately wide lateral projections on upper
beak; tooth-rows 4⁄6; upper rows subequal in length; fourth row interrupted
medially; length of lower rows 1-4 equal to upper rows; fifth and sixth lower
rows shorter; first lower row interrupted medially.
Body brown above; tip of snout brown; venter creamy gray; caudal musculature
creamy tan; caudal fin transparent; cream-colored crescent-shaped mark on
posterior edge of body; dark brown flecks on caudal musculature and all except
anterior two-thirds of ventral caudal fin; large brown square blotches on dorsum
of caudal fin; eye reddish brown in life.
Variation.—The variation in size and proportions is given in Table 2. With
the exception of one specimen having a short, broken, seventh tooth-row, all
specimens have 4⁄6 tooth-rows that are like those described above. In some
specimens the brown blotches on the dorsum of the caudal musculature are
fused and marked with cream-colored flecks.
Comparisons.—Ptychohyla leonhardschultzei differs from all other members
of the Ptychohyla euthysanota group in having a square snout, and further
differs from P. spinipollex in more numerous and smaller nuptial spines and in
transverse, instead of posteromedially slanting, vomerine processes between
the inner nares. Ptychohyla leonhardschultzei differs from P. euthysanota in
having a rostral keel and in having white flanks boldly spotted with black.
All small hylids that are sympatric with Ptychohyla leonhardschultzei are
either yellow (Hyla dendroscarta and H. melanomma) or green (Hyla erythromma,
which has a red eye, Hyla hazelae, which has a black line on the
canthus, and Ptychohyla ignicolor, which has red flash colors on the thighs).
Life History.—This frog has been found along streams in cloud
forests and in pine-oak forest. Males call from vegetation along
the stream or from rocks in and at the edge of the stream. The call
is a single, long, soft "wraack," repeated at intervals of anywhere
from several seconds to three or four minutes. Each note has a
duration of 0.62 to 0.95 of a second and a rate of 76 to 78 pulses
per second; the dominant frequency falls between 2700 and 2800
cycles per second (Pl. 11C).
Tadpoles were found in several streams in northern Oaxaca. A
tadpole having a total length of 21.1 mm. has three upper and four
lower tooth-rows well developed; the fourth upper and fifth lower
rows are weakly present, and the sixth lower row has not started to
develop. Two metamorphosed young have snout-vent lengths of
15.2 and 15.5 mm.
Remarks.—Four specific names have been applied to this species.
Ahl (1934:185) based his description of Hyla leonhardschultzei on
a small, poorly preserved female. Taylor (1944:41) proposed the
generic name Ptychohyla for a species (named therein as P. adipoventris)
of hylid having ventrolateral glands and horn-covered
nuptial spines. Obviously, Taylor was unaware that Hyla leonhardschultzei
was the same species. Earlier Taylor (1937:46) described
[Pg 327]
Hyla pinorum. The types of all of these species came from the
Pacific slopes of the Sierra del Sur in Guerrero. Examination of
the types and other available specimens shows that they are representatives
of a single species. The type of Hyla pinorum is an immature
male having a snout-vent length of 26.7 mm. All of these
specimens have the square snout and black and white flanks characteristic
of Ptychohyla leonhardschultzei. Although Shannon
(1951:473) based his description of Hyla milleri on a male having
well-developed ventrolateral glands, he overlooked the presence of
these glands in his description and discussion of relationships. The
acquisition of more specimens from northern Oaxaca has shown
that Hyla milleri is the same as Ptychohyla leonhardschultzei.
Distribution.—This species is known from pine-oak forest and cloud forest
on the Pacific slopes of the Sierra Madre del Sur in Guerrero and Oaxaca and
from the Atlantic slopes of the Sierra Madre Oriental in northern Oaxaca.
Specimens have been collected at elevations between 700 and 1650 meters.
Probably the species occurs in humid forests at similar elevations around the
eastern end of the Mexican Highlands in Oaxaca.
Specimens examined.—Mexico: Guerrero: Agua del Obispo, CNHM
123489-90, 126651, 106300, MCZ 29639, UIMNH 25047, 25049, USNM
114551; Malinaltepec, ZMB 34351, 34353. Oaxaca: 2.5 km. N of La Soledad,
KU 58061; San Lucas Camotlán, UIMNH 3201, USNM 123700-1; Vista Hermosa,
KU 64116-7, 64119, 68560 (tadpoles), 71344, 71717-8 (tadpoles),
UMMZ 119604; 5 km. S of Yetla, KU 60045 (tadpoles); 7.5 km. S of Yetla,
KU 64118, 68556-7 (tadpoles), 68559 (tadpoles), 68561 (2 young), 68630
(skeleton), UMMZ 115514-5, 118863 (tadpoles); 9 km. S of Yetla, KU 68558
(tadpoles).
Plate 15  Click to View Larger. | Ptychohyla spinipollex (Schmidt)
Hyla euthysanota, Dunn and Emlen, Proc. Acad. Nat. Sci. Philadelphia, 84:25, March 22, 1932. Hyla spinipollex Schmidt, Proc. Biol. Soc. Washington, 49:45-46, May 1, 1936 [Holotype.—MCZ 21300 from the mountains behind Ceiba, Depto. Atlantidad, Honduras; Raymond E. Stadelman collector]. Stuart, Misc. Publ. Mus. Zool. Univ. Michigan, 69:32-34, figs. 5-6, June 12, 1948; Contr. Lab. Vert. Biol. Univ. Michigan, 45:22, 52, 54, 57, May, 1950; Proc. Biol. Soc. Washington, 67:169, August 5, 1954. Ptychohyla spinipollex, Stuart, Contr. Lab. Vert. Biol. Univ. Michigan, 68:48, November, 1954. Duellman, Univ. Kansas Publ. Mus. Nat. Hist., 13:351, April 27, 1961. |
Diagnosis.—Rostral keel present; snout in lateral profile rounded above;
eyelid nearly as wide as interorbital region; flanks white with brown spots;
belly spotted; nuptial spines pointed and moderate in size.
Description.—The following description is based on KU 58054 from Finca
Los Alpes, Depto. Alta Verapaz, Guatemala (Pl. 15). Adult male having a
snout-vent length of 37.7 mm.; tibia length, 18.2 mm.; tibia length/snout-vent
length, 48.2 per cent; foot length, 15.8 mm.; head length, 11.7 mm.;
head length/snout-vent length, 31.0 per cent; head width, 11.7 mm.; head
width/snout-vent length, 31.0 per cent; diameter of eye, 3.6 mm.; diameter
of tympanum, 1.9 mm.; tympanum/eye, 52.7 per cent. Snout in lateral profile
[Pg 328]
nearly square, slightly rounded above, and in dorsal profile rounded with a
pointed tip resulting from vertical rostral keel; canthus pronounced; loreal
region barely concave; lips thick, rounded, and barely flaring; nostrils protuberant;
internarial distance, 3.0 mm.; top of head flat; interorbital distance,
3.7 mm., about equal to width of eyelid, 3.6 mm. A heavy dermal fold from
posterior corner of eye above tympanum to point above insertion of forearm,
covering upper edge of tympanum; tympanum round, its diameter equal to
its distance from eye. Forearm moderately robust, having faint dermal fold
on wrist; row of low, rounded tubercles along ventrolateral edge of forearm;
pollex only slightly enlarged, bearing triangular patch of moderate-sized,
pointed, horn-covered spines (38 on right, 43 on left); second finger noticeably
shorter than fourth; subarticular tubercles round, distal one on fourth
finger bifid; discs of fingers of moderate size, that of third finger slightly
smaller than diameter of tympanum; vestigial web between first and second
fingers; other fingers one-third webbed. Heels broadly overlapping when
hind limbs adpressed; tibiotarsal articulation reaches to middle of eye; distinct,
but low, tarsal fold extending length of tarsus; inner metatarsal tubercle
elevated, flat, and elliptical; outer metatarsal tubercle small and round, near
base of fourth toe; row of low indistinct tubercles from heel to base of fifth toe;
subarticular tubercles round; length of toes from shortest to longest 1-2-3-5-4,
the third and fifth being about equal in length; third and fifth toes webbed
to base of disc; fourth toe webbed to base of penultimate phalanx; discs of toes
slightly smaller than those on fingers. Anal opening near upper edge of
thighs; opening bordered laterally by moderately heavy dermal folds
and ventrolaterally by tubercles. Skin of dorsum and ventral surfaces of
forelimbs and shanks smooth; that of throat, belly, and ventral surfaces of
thighs granular. Ventrolateral glands barely evident. Tongue ovoid, barely
notched behind and slightly free posteriorly; vomerine teeth 2-3, situated
on ![]() -shaped elevations between round inner nares; openings to vocal sac
-shaped elevations between round inner nares; openings to vocal sac
large, one situated along inner posterior edge of each mandibular ramus.
Dorsal ground-color of head, body, and limbs grayish tan with dark brown
reticulations on head and body and dark brown transverse bars or spots on
limbs; first and second fingers cream color; third and fourth fingers and
webbing on hands pale grayish brown; anterior surfaces of thighs reddish tan;
posterior surfaces of thighs yellowish tan; tarsi and toes pale grayish tan with
brown flecks; webbing on foot pale brown; faint white stripe along ventrolateral
edges of tarsi and forearms; narrow white line above and beside anal
opening; no white stripe on edge of upper lip; axilla pale flesh-color; throat,
chest, and ventral surfaces of limbs pale creamy gray; belly white with
scattered brown flecks; flanks grayish white with dark brown flecks; ventral
surfaces of tarsi dark brown; ventrolateral glands grayish tan.
In life the dorsal ground-color of the head, body, fore limbs, and thighs
was yellowish tan (Pinkish Buff); dorsal surfaces of shanks and tarsi pale
yellowish tan (Pale Pinkish Buff); markings on head and back brown (Snuff
Brown) to dark brown (Chocolate); dark bands on limbs brown (Tawny-Olive);
first and second fingers creamy tan (Light Pinkish Cinnamon); posterior
surfaces of thighs creamy tan (Light Pinkish Cinnamon); third and fourth
fingers and webbing on hand grayish brown (Avellaneous); webbing on feet
dark brown (Olive Brown); axilla pale pink (Hydrangea Pink); flanks buff
[Pg 329]
(Cream-Buff) becoming yellow (Lemon Chrome) in groin; spots on flanks
dark brown (Clove Brown); iris dull grayish bronze (Orange-Citrine).
Variation.—The distal subarticular tubercle beneath the fourth finger is
bifid in about two-thirds of the specimens; in the rest it is round. The posterior
edge of the tongue varies from being emarginate to shallowly notched. In
most specimens the row of tubercles along the outer edge of the tarsus is
made up of discrete tubercles, but in some individuals the tubercles form a
nearly continuous dermal fold. Most specimens have the vomerine teeth
situated on ![]() -shaped elevations, but in some individuals the elevations are
-shaped elevations, but in some individuals the elevations are
more nearly transversely situated between the inner nares.
All 42 specimens from Finca Los Alpes, Guatemala, have dark brown
spots and flecks on the venter. Some individuals have only a few flecks on
the throat and a few large spots on the flanks, as does KU 64125. Other
specimens, such as KU 64132, have dense spotting over the entire venter.
The color of the dorsum varies from pale tan to dark brown with darker brown
markings; the white line above the anus is present in all specimens, but in
some it is indistinct. KU 58053 and 64127 have a dark brown dorsum with
large pale tan, square blotches; in life the blotches were pale tan (Pinkish
Buff); the rest of the dorsum was dark brown (Sayal Brown). KU 58052
is dark brown with many small white flecks on the dorsum; in life the dorsum
was deep olive brown (Dark Olive).
Aside from the differences mentioned above, all specimens from Guatemala
are similar in coloration. Three specimens from Honduras (MCZ 21300 and
UMMZ 113102-3) have unspotted white venters. MCZ 21300, the holotype
of P. spinipollex, lacks a white stripe above the anal opening, whereas the
stripe is indistinct in UMMZ 113102-3.
Description of tadpole.—The following description is based on KU 60053
from Fina Los Alpes, Depto. Alta Verapaz, Guatemala (Figs. 4D and 6D).
No limb buds; total length, 37.2 mm.; body length, 12.2 mm.; body length/total
length, 32.8 per cent. Body rounded, not depressed, as wide as deep, ovoid
in dorsal profile; mouth directed ventrally; eyes small, directed dorsolaterally;
nostrils barely protuberant and directed anterolaterally, somewhat closer to eye
than snout; spiracle sinistral and posteroventrad to eye; anal tube dextral.
Caudal fin low, bluntly rounded posteriorly; greatest depth of caudal musculature
about one-half depth of caudal fin; musculature extends nearly to tip of
tail.
Mouth large; lips having deep lateral folds; two complete rows of papillae
on lips; six or seven rows of papillae laterally; beaks moderately developed,
bearing fine blunt serrations; slender lateral projections on upper beak; tooth-rows
4⁄7; upper rows subequal in length; fourth row interrupted medially; lower
rows 1-4 equal in length to upper rows; lower rows 5-7 decreasing in length;
first lower row interrupted medially.
Top of head and tip of snout brown; venter creamy gray; caudal musculature
tan; caudal fin transparent; faint cream-colored, narrow, crescent-shaped
mark on posterior edge of body, not bordered posteriorly by dark brown mark;
brown flecks scattered on caudal musculature and caudal fin; only a few flecks
on anterior half of ventral caudal fin; eye bronze-color in life.
Variation.—The variation in size and proportions as compared with tadpoles
of other species is given in Table 2. Of the 57 tadpoles of this species that I
[Pg 330]
have examined, 21 have only six lower tooth-rows, although in some of these
specimens a faint ridge for a seventh row is present. In those specimens having
seven lower rows, the seventh often is broken.
There is considerable variation in coloration. None has a distinct cream-colored,
crescent-shaped mark bordered posteriorly by a dark brown bar or
triangle, as in the other species in the Ptychohyla euthysanota group. Most
specimens have a rather indistinct crescent; some have no crescent. In a few
specimens there is a weakly outlined dark mark posterior to the crescent.
Some specimens in a series of tadpoles from 32 kilometers north of Morazán,
Baja Verapaz, Guatemala, have faint dorsal blotches on the dorsal musculature,
much like those in tadpoles of Ptychohyla leonhardschultzei.
Comparisons.—Ptychohyla spinipollex differs from all other species in the
genus by having moderate-sized, instead of small, pointed nuptial spines; also
it has fewer spines than the other species (see discussion of this character in
Analysis of Data). The nearly equal interorbital breadth and width of the
upper eyelid also is diagnostic of this species.
Other hylids sympatric with Ptychohyla spinipollex include three species of
Plectrohyla, each of which has a bony prepollex, heavy body, and rugose skin
on the dorsum. The only other sympatric hylid that could be confused with
Ptychohyla spinipollex is Hyla bromeliacea, which has a round snout and
yellowish tan dorsum not marked with dark brown.
Life History.—At Finca Los Alpes, Guatemala, in July, 1960, and
in August, 1961, calling males were found on bushes and trees along
cascading mountain streams. The breeding call consists of a soft
"wraack," repeated at intervals of 45 seconds to four minutes. Each
note has a duration of about .46 of a second and a rate of 147 pulses
per second. The dominant frequency is 4300 cycles per second
(Pl. 11A).
Tadpoles have been found in cascading mountain streams. Two
metamorphosed young have snout-vent lengths of 15.0 and 15.5 mm.
Remarks.—There is little doubt that all of the specimens herein
referred to Ptychohyla spinipollex are conspecific. However, the
three specimens from Honduras, including the type of Ptychohyla
spinipollex, differ from Guatemalan specimens in lacking all dark
spotting on the venter. Additional specimens from Honduras and
eastern Guatemala may show that two subspecies are recognizable,
in which case the nominal subspecies will be the population in
Honduras.
Distribution.—This species lives in cloud forests at elevations of 800 to
1700 meters on the Atlantic side of the Guatemalan Highlands from the Sierra
de Cuchumatanes in western Guatemala eastward to central Honduras.
Specimens examined.—Guatemala: Alta Verapaz: Finca Chichén, UMMZ
90876 (tadpoles); Finca Los Alpes, KU 58052-60, 59939 (skeleton), 60053
(tadpoles), 64122-41, 68562, 68563 (tadpoles), 68631-2 (skeletons), MCZ
35000-1, UMMZ 90873, 90874 (3), 90875 (tadpoles); La Primavera, UMMZ
90877 (tadpoles); Panzamalá, UMMZ 90878 (tadpoles). Baja Verapaz: 32
[Pg 331]
km. N of Morazán, KU 68564 (tadpoles); Santa Elena, UMMZ 98119, 98120
(2). Huehuetenango: 1 km. E of Barillas, UMMZ 123136-7 (tadpoles).
Progreso: Finca Bucaral, UMMZ 106783 (3), 123138 (tadpoles), S-1292
(skeleton).
Honduras: Atlantidad: Mountains behind Ceiba, MCZ 21300. Morazán:
Cerro Uyuca, UMMZ 123102-3.
Two species in group; adults having only vestige of web between fingers
and lacking tarsal fold; pollex of breeding males lacking spinous, horny, nuptial
tuberosities; mouth of tadpole greatly expanded, funnel-shaped, lacking lateral
folds, and having 3⁄3 tooth-rows; breeding call consisting of series of short notes.
Diagnosis.—Diameter of tympanum more than half of diameter of eye;
internarial region depressed; toes three-fourths webbed; no red flash-colors on
thighs.
Plate 16 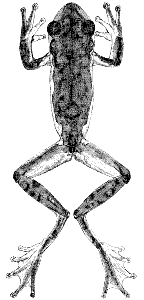 Click to View Larger. | Ptychohyla schmidtorum schmidtorum Stuart Ptychohyla schmidtorum Stuart, Proc. Biol. Soc. Washington, 67:169-172, August 5, 1954 [Holotype.—CNHM 27055 from El Porvenir (17 kilometers air-line west of San Marcos), Depto. San Marcos, Guatemala; Karl P. Schmidt collector]. Duellman, Univ. Kansas Publ. Mus. Nat. Hist., 13:351, 355, April 27, 1961. |
Diagnosis.—Vomerine teeth 5-11; dorsum dark brown; white spot below
eye; eye red in life.
Description.—The following description is based on KU 58043 from Finca
La Paz, Depto. San Marcos, Guatemala (Pl. 16). Adult male having snout-vent
length of 31.6 mm.; tibia length, 15.0 mm.; tibia length/snout-vent length,
47.5 per cent; foot length, 12.5 mm.; head length, 10.2 mm.; head length/snout-vent
length, 32.3 per cent; head width, 9.9 mm.; head width/snout-vent length,
31.3 per cent; diameter of eye, 3.4 mm.; diameter of tympanum, 1.8 mm.;
tympanum/eye, 52.9 per cent. Snout in lateral profile nearly square, slightly
rounded above and below, and in dorsal profile bluntly squared; canthus pronounced;
loreal region concave; lips thick, rounded, and flaring; nostrils protuberant;
internarial distance, 2.2 mm.; internarial region depressed; top of
head flat; interorbital distance, 3.4 mm., much greater than width of eyelid,
2.5 mm. Thin dermal fold from posterior corner of eye above tympanum to
insertion of forelimb, covering upper edge of tympanum; tympanum round,
its diameter equal to its distance from eye. Forearm slender, lacking distinct
dermal fold on wrist; row of low rounded tubercles along ventrolateral edge
of forearm; pollex slightly enlarged; no nuptial spines; second and fourth
fingers about equal in length; subarticular tubercles small and round, distal
one beneath fourth finger bifid; discs small, that of third finger noticeably
smaller than tympanum; no web between first and second fingers; vestige of
web between other fingers. Heels overlap when hind limbs adpressed;
tibiotarsal articulation reaches to middle of eye; no tarsal fold; inner metatarsal
tubercle large, flat, and elliptical; outer metatarsal tubercle small, ovoid,
slightly more distal than inner; subarticular tubercles round; length of digits
from shortest to longest 1-2-5-3-4; third and fifth toes webbed to base of
discs; fourth toe webbed to base of penultimate phalanx; discs of toes smaller
[Pg 332]
than on fingers. Anal opening directed posteriorly at upper edge of thighs;
no anal flap; pair of large tubercles below anal opening and smaller tubercles
farther below. Skin of dorsum and ventral surfaces of forelimbs and shanks
smooth; that of belly and ventral surfaces of thighs granular. Ventrolateral
glands well developed, reaching axilla and groin and narrowly separated on
chest. Tongue ovoid, emarginate posteriorly, and only slightly free behind;
vomerine teeth 3-3, situated on small triangular elevations between ovid inner
nares; openings to vocal sac large, one situated along inner posterior edge of
each mandibular ramus.
Dorsum of head, body, and limbs reddish brown with indistinct, irregular
darker brown markings on body and dark brown transverse bands or spots
on limbs; first and second fingers creamy white; third and fourth fingers brown;
dorsal surfaces of tarsi and third, fourth, and fifth toes tan with brown spots;
first and second toes and webbing on feet creamy tan; enamel-white stripe
along edge of upper lip continuing over, and on posterior edge of, forearm to
groin, expanded to form spot below eye; belly white, unspotted; ventrolateral
glands pale brown; ventral surfaces of hind limbs and anterior and posterior
surfaces of thighs cream color; enamel-white stripe on heel; creamy white stripe
along ventrolateral edges of tarsi and forearms.
In life dorsum reddish brown (Terra Cotta) with dark brown (Burnt
Umber) markings; first and second fingers and first and second toes orange-yellow
(Light Orange-Yellow); posterior surfaces of thighs pale reddish tan
(Ochraceous-Salmon); webbing on feet yellowish tan (Deep Colonial Buff);
belly white; iris red (Nopal Red).
Variation.—Little variation in structural characters was observed. All but
five specimens have bifid subarticular tubercles beneath the fourth finger.
Three specimens have cordiform tongues, and in four others the tongue is
ovoid and shallowly notched behind; all other specimens have an emarginate
ovoid tongue.
Some individuals when active at night had a pale brown (Ochraceous-Tawny)
dorsum with dull olive green (Dark Olive Buff) markings. Otherwise
there was no noticeable variation in color.
Description of tadpole.—The following description is based on KU 60051
from Finca La Paz, Depto. San Marcos, Guatemala (Figs. 5A and 6E). Small
hind limbs; total length, 37.9 mm.; body length, 11.6 mm.; body length/total
length, 30.6 per cent. Body only slightly depressed, nearly as deep as wide,
in dorsal profile ovoid, widest just posterior to eyes; in lateral profile snout
rounded; mouth directed ventrally; eyes small, directed dorsolaterally; nostrils
barely protuberant, directed anteriorly, somewhat closer to eye than snout;
spiracle sinistral and posteroventrad to eye; anal tube dextral. Tail long and
slender; caudal fin low and rounded posteriorly; depth of caudal musculature
one-half greatest depth of caudal fin; musculature not extending to tip of tail.
Mouth large; thin fleshy lips greatly expanded and forming large funnel-shaped
disc; width of mouth two-thirds greatest width of body; outer edge
of lips having one row of small papillae; inner surfaces of mouth smooth;
scattered large papillae forming one nearly complete row around teeth; other
large papillae laterally; beaks moderately developed, bearing long, pointed
denticulations; no lateral projections on upper beak; tooth-rows 3⁄3, all short;
second and third upper rows subequal in length; first upper row shorter; first
[Pg 333]
and third upper rows interrupted medially; first lower row interrupted medially,
equal in length to second and third upper rows; second lower row slightly
shorter; third lower row shortest.
Body mottled brown and creamy gray above and below; mouth colored
like body; caudal musculature creamy tan; caudal fin transparent; dark brown
streak mid-laterally on anterior third of caudal musculature; rest of tail and
all of caudal fin heavily flecked with brown; eye red in life.
Variation.—The third upper tooth-row is interrupted in all specimens; in
some individuals the first upper and first lower rows are complete. The variation
in size and proportions is given in Table 2. The dark brown lateral
streak on the anterior part of the caudal musculature is distinct on most specimens;
the only other variation in coloration is in the amount of brown flecking
on the caudal musculature and fin.
Comparisons.—Ptychohyla schmidtorum schmidtorum differs from P. schmidtorum
chamulae as stated in the diagnosis and in having pale creamy tan, as
opposed to dark brown, webbing on the feet; and from P. ignicolor in having
a depressed, as opposed to a flat, internarial region. Tadpoles of P. s. schmidtorum
have a mottled appearance, as opposed to the more uniform brown color
of P. s. chamulae.
Ptychohyla schmidtorum schmidtorum and several species of Plectrohyla
are sympatric. All species of the latter genus have a bony prepollex, rugose
skin on the dorsum, and heavy body; also sympatric is Ptychohyla e. euthysanota,
which has a tarsal fold and in breeding males spinous nuptial tuberosities.
Life History.—This species breeds in clear mountain streams
where males call from vegetation along the stream. The call consists
of series of short notes, three to nine notes per series, sounding
like "raa-raa-raa." The duration of each note is approximately .065
of a second, and has a rate of 96 to 119 pulses per second; the
dominant frequency is about 3400 cycles per second. The call is
almost indistinguishable from that of Ptychohyla schmidtorum
chamulae.
Tadpoles and metamorphosing young were found at Finca La
Paz, Guatemala, in late July, 1960. Two young lacking tails but
not having completely developed mouths have snout-vent lengths
of 14.2 and 14.6 mm. L. C. Stuart collected four metamorphosing
young at Finca La Paz on May 6, 1949. By May 10 the frogs were
completely metamorphosed, at which time they had snout-vent
lengths of 15.5 to 17.0 (average 16.1) mm.
Remarks.—There is no doubt that this frog is most closely related
to Ptychohyla schmidtorum chamulae, even though the ranges
of the two subspecies are separated by the interior depression of
Chiapas. Since at least at Finca La Paz, Guatemala, P. s. schmidtorum
occurs with P. e. euthysanota, it is surprising that the former
species has not been found at more localities along the Pacific slopes
[Pg 334]
on northern Central America. At Finca La Paz in July, 1960, P. s.
schmidtorum was more abundant than P. e. euthysanota.
Distribution.—This species is known only from a limited area at elevations
between 1300 and 2200 meters on the Pacific slopes of the Sierra Madre
in extreme eastern Chiapas and western Guatemala.
Specimens examined.—Mexico: Chiapas: Finca Irlandia, UMMZ 105429-30.
Guatemala: San Marcos: El Porvenir, CNHM 20755, 20761, 69904,
UMMZ 80918; Finca La Paz, 2 km. W of La Reforma, KU 58016-44, 59940-2
(skeletons), 60050 (3 young), 60051 (tadpoles), 60052 (4 young), MCZ
34998-9, UMMZ 123144-7 (tadpoles).
Plate 17 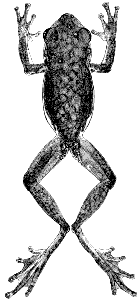 Click to View Larger. | Ptychohyla schmidtorum chamulae Duellman Ptychohyla chamulae Duellman, Univ. Kansas Publ. Mus. Nat. Hist., 13: 354-357, pl. 25, fig. 2, April 27, 1961 [Holotype.—KU 58063 from 6.2 kilometers south of Rayón Mescalapa, Chiapas, México; William E. Duellman collector]. |
Diagnosis.—Vomerine teeth 4-6; dorsum bright green; white lateral stripe;
eye reddish bronze in life.
Description.—The following description is based on KU 58069 from 6.2
kilometers south of Rayón Mescalapa, Chiapas, México (Pl. 17). Adult male
having a snout-vent length of 27.6 mm.; tibia length, 13.0 mm.; tibia
length/snout-vent length, 47.1 per cent; foot length, 10.8 mm.; head length,
9.6 mm.; head length/snout-vent length, 34.7 per cent; head width, 9.2 mm.;
head width/snout-vent length, 33.1 per cent; diameter of eye, 3.0 mm.;
diameter of tympanum, 1.6 mm.; tympanum/eye, 53.3 per cent. Snout in
lateral profile nearly square, slightly rounded above and below, and in dorsal
profile blunt, almost square; canthus pronounced; loreal region concave; lips
thick, rounded and flaring; nostrils protuberant; internarial distance, 2.5 mm.; internarial
region slightly depressed; top of head flat; interorbital distance, 3.3
mm., much greater than width of eyelid, 2.5 mm. Thin dermal fold, from
posterior corner of eye above tympanum to insertion of fore limb, covering
upper edge of tympanum; tympanum nearly round, its diameter equal to its
distance from eye. Forearm slender, lacking distinct fold on wrist; row of
low, rounded tubercles on ventrolateral surface of forearm; pollex slightly enlarged,
without nuptial spines; second and fourth fingers equal in length; subarticular
tubercles round, that under fourth finger bifid; discs small, that of
third finger noticeably smaller than tympanum; no web between first and
second fingers; vestige of web between other fingers. Heels overlapping when
hind limbs adpressed; tibiotarsal articulation reaches to middle of eye; no
tarsal fold; inner metatarsal tubercle large, flat, and elliptical; outer metatarsal
tubercle small, elliptical, slightly more distal than inner; subarticular tubercles
round; length of digits from shortest to longest 1-2-5-3-4; third and fifth toes
webbed to base of disc; fourth toe webbed to base of penultimate phalanx;
discs smaller on toes than on fingers. Anal opening directed posteriorly at
upper edge of thighs; no anal flap; pair of large tubercles below anal opening.
Skin of dorsum and of ventral surfaces of forelimbs and shanks smooth; that
of throat, belly, and ventral surfaces of thighs granular. Ventrolateral glands
well developed, not reaching axilla or groin and broadly separated midventrally.
Tongue cordiform, shallowly notched behind and only slightly free
posteriorly; vomerine teeth 2-3, situated on small triangular elevations
[Pg 335]
between ovoid inner nares; openings to vocal sac large, one situated along inner
posterior edge of each mandibular ramus.
Dorsum of head, body and limbs reddish brown with dark purplish brown
markings on back and shanks; first finger creamy tan; other fingers pale
brown; dorsal surfaces of tarsi, third, fourth, and fifth toes dull tan with
brown spots; first and second toes creamy tan; webbing on feet brown;
anterior and posterior surfaces of thighs tan; faint creamy white stripe along
ventrolateral edges of tarsi and forearms; enamel-white stripe on heel; axilla
and groin gray; enamel-white stripe on edge of upper lip, continuing onto
proximal upper surfaces of forelimb and on flanks to groin, widened under eye
to form large spot, and bordered below on flanks by dark brown stripe; white
stripe above and white spots below anal opening; throat and chest white;
belly and ventral surfaces of limbs cream color; brown dash on either side of
chin and brown spot on throat near angle of jaws; few brown flecks on belly;
ventrolateral glands orange-tan; ventral surfaces of tarsi and feet brown.
In life, dorsal surfaces of head, body, and limbs bright green (Shamrock
Green); first and second fingers pale orange (Apricot Yellow); stripe on upper
lip and spot below eye enamel-white; stripe on flanks silvery white, bordered
below by brown (Saccardo's Umber) brown; anterior and posterior surfaces of
thighs yellowish brown (Old Gold); webbing of feet dull brown (Brownish
Olive); belly deep yellow (Amber Yellow); iris reddish bronze (English Red).
Variation.—Tubercles beneath the fourth fingers are bifid in 20 specimens
and rounded in all others. The tongue is emarginate in 12 specimens and
cordiform in all others. In most specimens the white stripe on the upper lip
continues onto the flanks and to the groin; in five specimens the stripe terminates
above the forearm, and in three it terminates at mid-flank. The lateral
stripe is absent in two specimens. All specimens were uniform green above
when found at night; later some changed to pale green (Light Oriental Green)
on the dorsum with irregular yellowish tan (Naples Yellow) blotches. Most
males have brown flecks on the throat and ventrolateral gland, but some
specimens are immaculate below, and one has dark brown mottling on the
throat. Several males have a round, orange-tan glandular area on the chin,
as does P. ignicolor.
Description of tadpole.—The following description is based on KU 58199
from 6.2 kilometers south of Rayón Mescalapa, Chiapas, México (Figs. 5B and 6F). Hind limbs small; total length, 39.0 mm.; body length, 10.5 mm.;
body length/total length, 26.9 per cent. Body barely depressed, only slightly
wider than deep, widest just posterior to eyes; in dorsal profile ovoid; mouth
directed ventrally; eyes small, directed dorsolaterally; nostrils barely protuberant,
directed anterodorsally, slightly closer to eye than snout; spiracle
sinistral and posteroventrad to eye; anal tube dextral. Tail long and slender;
caudal fin low, rounded posteriorly; depth of caudal musculature one-half
greatest depth of caudal fin; musculature not extending to tip of tail.
Mouth large; thin fleshy lips greatly expanded and forming funnel-shaped
disc; outer edge of lips having one row of small papillae; inner surfaces of
mouth smooth; scattered large papillae forming nearly one complete row
around teeth; other papillae laterally; beaks moderately developed, bearing
long, pointed denticulations; no lateral projections on upper beak; tooth-rows
3⁄3, all short; second and third upper rows subequal in length; first upper row
shorter; first and third upper rows interrupted medially; first lower row
[Pg 336]
interrupted medially, equal in length to second and third upper rows; second lower
row slightly shorter; third lower row shortest.
Body dark brown above and dark gray below; fleshy part of mouth creamy
gray mottled with dark brown; caudal musculature pale tan with heavy suffusion
of brown flecks; caudal fin transparent with brown spots; dark brown
streak mid-laterally on anterior one-fifth of caudal musculature, bordered below
by cream-colored spot; eye brown in life.
Variation.—The third upper tooth-row is interrupted in all specimens, but
in some individuals the first upper row and first lower row are complete. The
only noted variation in color is the intensity of brown pigmentation on the
caudal musculature, which in most specimens is sufficiently dense to make the
tail look brown. In some specimens the mid-lateral streak is indistinct, and
the pale spot below the streak is absent.
Comparisons.—Aside from the characters listed in the diagnosis, Ptychohyla
schmidtorum chamulae differs from P. schmidtorum schmidtorum by having
dark brown webbing on the feet, instead of pale creamy tan webbing, and in
having in life a yellow venter, instead of a white venter. Ptychohyla ignicolor
also is green in life, but has red flash-colors on the thighs, red webbing on
the feet, and lacks the white lateral stripe diagnostic of P. schmidtorum
chamulae.
Plectrohyla matudai matudai and P. guatemalensis are sympatric with
Ptychohyla schmidtorum chamulae. Each of the first two has a bony prepollex,
rugose skin on the dorsum, and heavy body. Also living with Ptychohyla
chamulae are Hyla chaneque, a large species having a tuberculate dorsum and
webbed fingers, and Hyla bivocata, a small yellow species having a broad, flat
head, small indistinct tympanum, and an axillary membrane.
Life History.—Calling males were found on leaves of herbs and
bushes by cascading streams in cloud forest. The call consists of
series of short notes, three to nine notes per series, sounding like
"raa-raa-raa." The duration of each note is .054 to .070 of a second,
and has a rate of 96 to 110 pulses per second. The dominant
frequency falls between 3350 and 3450 cycles per second (Pl. 11D).
The call is almost indistinguishable from that of Ptychohyla schmidtorum
schmidtorum.
Tadpoles were found in the cascading streams; the smallest tadpole
has a total length of 17.2 mm. and has only 3⁄2 tooth-rows. At a
stream 6.2 kilometers south of Rayón Mescalapa, Chiapas, metamorphosing
young were found on June 16 and August 5, 1960.
Each of two completely metamorphosed young has a snout-vent
length of 15.7 mm. Another having a snout-vent length of 16.2 mm.
has a tail stub 2 mm. long and a completely metamorphosed mouth.
Two others have snout-vent lengths of 13.6 and 14.1 mm. and tail
lengths of 11.5 and 8.1 mm. respectively; in these the mouth parts
are incompletely metamorphosed.
Remarks.—Duellman (1961:354) described Ptychohyla chamulae
[Pg 337]
and stated that it probably was most closely related to P. schmidtorum.
Further study has revealed additional resemblance in morphological
and behavioral details. It is concluded that the two
populations are more realistically treated as subspecies than as
species. The geographic ranges, as now known, are disjunct.
Ptychohyla schmidtorum chamulae inhabits cloud forest on the
Atlantic slopes of the Chiapan Highlands, whereas P. s. schmidtorum
lives in cloud forest on the Pacific slopes of the Sierra Madre
in Chiapas and Guatemala. Between their known geographic
ranges are the pine clad Sierra Madre and Chiapan Highlands, and
intervening sub-humid Grijalva Valley.
Distribution.—This species is known only from elevations between 1500 and
1700 meters on the Atlantic slopes of the Chiapan Highlands; it is to be expected
in cloud forests on the northern slopes of the Sierra de Cuchumatanes
in Guatemala.
Specimens examined.—Mexico: Chiapas: 15 km. N of Pueblo
Nuevo Solistahuacán, UMMZ 123325 (4); 16.5 km. N of Pueblo Nuevo
Solistahuacán, UMMZ 123322 (10); 18 km. N of Pueblo Nuevo Solistahuacán,
UMMZ 121395-9, 123324 (8), 123326 (5); 18.6 km. N of Pueblo Nuevo
Solistahuacán, UMMZ 123323 (4); 5.6 km. S of Rayón Mescalapa, KU 58062,
58200 (tadpoles); 6.2 km. S of Rayón Mescalapa, KU 58063-74, 58199 (tadpole),
58234-8, 59936 (skeleton).
Plate 18 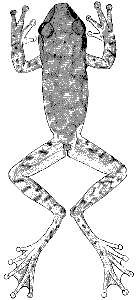 Click to View Larger. | Ptychohyla ignicolor Duellman Ptychohyla ignicolor Duellman, Uni. Kansas Publ. Mus. Nat. Hist., 13:352-353, pl. 25, fig. 1, April 27, 1961 [Holotype.—UMMZ 119603 from 6 kilometers south of Vista Hermosa, Oaxaca, México; Thomas E. Moore collector]. |
Diagnosis.—Diameter of tympanum less than one-half diameter of eye;
internarial region flat; 3-7 vomerine teeth; toes one-half webbed; no white
spot below eye; no lateral white stripe; in life dorsum green; groin and thighs
having bright red flash-colors.
Description.—The following description is based on UMMZ 119603 from
6 kilometers south of Vista Hermosa, Oaxaca, México (Pl. 18). Adult male
having a snout-vent length of 30.0 mm.; tibia length, 14.6 mm.; tibia length/snout-vent
length, 48.7 per cent; foot length, 12.3 mm.; head length, 9.2 mm.;
head length/snout-vent length, 30.7 per cent; head width, 9.3 mm.; head
width/snout-vent length, 31.0 per cent; diameter of eye, 3.2 mm.; diameter
of tympanum, 1.3 mm.; tympanum/eye, 40.6 per cent. Snout in lateral profile
square, and in dorsal profile rounded; canthus pronounced; loreal region
slightly concave; lips moderately flaring; top of head flat; nostrils protuberant;
internarial distance, 2.8 mm.; internarial region flat; interorbital distance,
3.3 mm., much broader than width of eyelid, 2.8 mm. A heavy dermal fold
from posterior corner of eye above tympanum to insertion of forelimb, covering
upper edge of tympanum; tympanum elliptical, its greatest diameter equal
to its distance from eye. Forearm moderately robust having distinct dermal
fold on wrist; pollex moderately enlarged without nuptial spines; second and
fourth fingers equal in length; subarticular tubercles round, none is bifid;
[Pg 338]
discs on fingers moderate in size, that on third finger slightly larger than
tympanum; no web between first and second fingers; vestige of web between
other fingers. Heels overlap when hind limbs adpressed; tibiotarsal articulation
extends to anterior corner of eye; no tarsal fold; inner metatarsal tubercle
large, flat, and elliptical; outer metatarsal tubercle near inner one and triangular
in shape; subarticular tubercles round; length of digits from shortest
to longest 1-2-5-3-4; third toe webbed to proximal end of penultimate phalanx;
fourth toe webbed to distal part of antepenultimate phalanx; fifth toe webbed
to middle of penultimate phalanx; discs on toes smaller than on fingers. Anal
opening directed posteriorly at upper edge of thighs; no anal flap; pair of
large tubercles below anal opening; small tubercles ventral and lateral to these.
Skin of dorsum and ventral surfaces of limbs smooth; that of throat and belly
granular. Ventrolateral glands noticeably thickened, extending from axilla
nearly to groin and only narrowly separated midventrally on chest; skin of
anterior part of chin thickened and glandular. Tongue cordiform, shallowly
notched behind and only slightly free posteriorly; vomerine teeth 0-3, situated
on rounded elevations between somewhat larger, round inner nares; openings
to vocal sac large, one situated along posterior margin of each mandibular
ramus.
Dorsal ground-color of head, body, and limbs dull brown with dark brown
reticulations on head and body and dark brown transverse bands or spots on
limbs; first and second fingers cream color; third and fourth fingers dull tan;
anterior surfaces of thighs pale brown; posterior surfaces of thighs cream
color with heavy suffusion of brown; dorsal surfaces of tarsi and third, fourth,
and fifth toes dull brown with dark brown spots; first and second toes creamy
white; webbing on foot brown; axilla and groin cream color; flanks brown; no
white stripes on edge of upper lip or on flank; faint, barely discernible tan
streak above anal opening; faint creamy tan line on ventrolateral edges of
tarsi; throat, belly, ventral surfaces of limbs, inner edges of tarsi, and first
toes cream color; outer ventral surfaces of tarsi and other toes brown; chest
and throat spotted with brown; ventrolateral and chin glands orange-brown.
In life the dorsum was uniform green (Cosse Green) becoming paler green
(Bright Green-Yellow) on flanks, later changing to paler green (Javel Green)
on dorsum with irregular darker green (Lettuce Green) markings and greenish
yellow (Green-Yellow) on flanks; anterior and posterior surfaces of thighs,
ventral surfaces of shanks, anterior surfaces of tarsi, and upper proximal surfaces
of first, second, and third toes red (Coral Red); venter pale creamy
yellow (Sulfur Yellow); iris pale golden color (Aniline Yellow).
Variation.—Of 13 specimens, six have a cordiform tongue; the others have
an emarginate tongue. Five specimens have round subarticular tubercles
beneath the fourth fingers; six specimens have a bifid tubercle on one hand,
and two specimens have bifid tubercles on both hands. A round gland is
present on the chin of all specimens; in some the gland is barely visible, but
in others it is large and distinct. In two specimens the ventrolateral glands
are weakly developed; in the others the glands are well developed and orange-tan.
The white anal stripe varies from a thin line to a series of white flecks.
Dark brown or black flecks are present on the throat, chest, and flanks of all
specimens. In some the flecks are small and widely scattered; in others the
flecks are larger and more numerous. All specimens were pale green above
[Pg 339]
when calling at night; later they changed to dull green with darker green
reticulations. The flash-colors on the thighs and in the groin vary from red
to orange-red or brownish red.
Description of tadpole.—The following description is based on KU 71716
from Vista Hermosa, Oaxaca, México (Figs. 5C and 6G). Hind limbs small;
total length, 39.6 mm.; body length, 11.8 mm.; body length/total length, 29.8
per cent. Body moderately depressed, only slightly wider than deep, in dorsal
profile ovoid, widest just posterior to eyes; in lateral profile snout rounded;
mouth directed ventrally; eyes small, directed dorsolaterally; nostrils barely
protuberant, directed anteriorly, somewhat closer to eye than snout; spiracle
sinistral and posteroventrad to eye; anal tube dextral. Tail long and slender;
caudal fin low and rounded posteriorly; depth of caudal musculature about
one-half greatest depth of caudal fin; musculature not extending to tip of
tail.
Mouth large; thin fleshy lips greatly expanded and forming large funnel-shaped
disc; width of mouth about two-thirds greatest width of body; outer
edge of lips having one row of small papillae; inner surface of mouth smooth;
scattered large papillae forming one nearly complete row around teeth; other
papillae laterally; one large papilla just above each end of first lower tooth-row;
beaks moderately developed bearing long, pointed denticulations; no
lateral projections on upper beak; tooth-rows 3⁄3, all short; second and third
upper rows subequal in length; first upper row shorter; first lower row equal
in length to second and third upper rows; second lower row slightly shorter;
third lower row shortest.
Body creamy gray with dark brown flecks above and below; mouth colored
like body; caudal musculature creamy tan; caudal fin transparent; dark brown
streak on anterior third of caudal musculature; rest of tail and all of caudal fin,
except anterior two-thirds of ventral fin, heavily flecked with brown; iris silvery
bronze color in life.
Variation.—The only other known tadpole, which was collected with the
individual described above, differs in having only two upper tooth-rows. The
first upper tooth-row seems not to have developed.
Comparisons.—From P. schmidtorum chamulae and P. s. schmidtorum, P.
ignicolor differs as follows: Tympanum smaller; snout more nearly square; less
webbing on toes; internarial region flat instead of depressed; white lateral
stripes lacking.
Ptychohyla ignicolor and several small and moderate sized hylids are
sympatric. From P. ignicolor these hylids can be distinguished as follows:
Hyla dendroscarta has a round snout and yellow dorsum; Hyla erythromma
has a round snout, green dorsum, white flanks, and a red eye; Hyla hazelae has
a tarsal fold, green dorsum, and a black line on the canthus; and Ptychohyla
leonhardschultzei has a tarsal fold, brown dorsum, black and white flanks, and
horny nuptial spines in breeding males.
Life History.—At Vista Hermosa, Oaxaca, males were calling on
vegetation above small streams on March 30, 1959, and on June 28,
1962; males were found on vegetation overhanging a stream 6 kilometers
south of Vista Hermosa on June 27 and July 3, 1962. The
call consists of a series of short notes, three to thirteen notes per
[Pg 340]
series, sounding like "raa-raa-raa." The duration of each note is
about .08 of a second and has a rate of 123 to 129 pulses per second.
The dominant frequency of notes in short series is about 2100 cycles
per second, whereas the dominant frequency of notes in long series
is about 3150 cycles per second (Pl. 11E).
On June 28, 1962, two tadpoles of this species were found in a
quiet pool in a spring-fed rivulet at Vista Hermosa, Oaxaca. Females
are unknown.
Remarks.—The absence of a tarsal fold and of nuptial spines in
breeding males, the nature of the breeding call, and the form of
tadpole are characters that place Ptychohyla ignicolor in the P.
schmidtorum-group.
Distribution.—This species is known from only two localities at elevations
of 1500 and 1850 meters in the cloud forest on the northern (Atlantic) slopes
of the Sierra Madre Oriental in northern Oaxaca.
Specimens examined.—Mexico: Oaxaca: Vista Hermosa, KU 71334, 71716
(tadpoles), UMMZ 119602; 6 km. S of Vista Hermosa, KU 71335-42, 71343
(skeleton), UMMZ 119603, 123327 (2).
The distribution of species of Ptychohyla reflects the distribution
of cloud forest in southern México and northern Central America.
The frogs are restricted to mountainous areas, usually at elevations
higher than 1000 meters above sea level. Ptychohyla does not range
to great heights in the mountains, where west of the Isthmus of
Tehuantepec the mountain streams are inhabited by frogs of the
Hyla bistincta group, and in Chiapas and Guatemala by species of
Plectrohyla.
Frogs of the Ptychohyla euthysanota group have a greater combined
geographic range than the species comprising the Ptychohyla
schmidtorum group (Fig. 7). No two species in the same group
are sympatric, but members of different groups are sympatric in at
least parts of their ranges. Apparently P. leonhardschultzei ranges
around the southern edge of the Mexican Highlands, where the
species occurs on both Atlantic and Pacific slopes; as can be seen
from the distribution map, there are many gaps in the known range
of this species. The range of P. euthysanota euthysanota is along
the Pacific slopes of the Sierra Madre in Chiapas, Guatemala, and
El Salvador, whereas that of P. euthysanota macrotympanum is
along the southern interior slopes of the Central Highlands of
Chiapas and the Sierra de Cuchumatanes in Guatemala. Ptychohyla
[Pg 341]
spinipollex occurs on the wet Atlantic slopes of the Guatemalan and
Honduranean Highlands; the range of the species in Honduras is
poorly known.

Fig. 7. Map showing locality records for the species and subspecies of
Ptychohyla.
The frogs of the Ptychohyla schmidtorum group have more restricted
geographic ranges than members of the former group.
Ptychohyla schmidtorum schmidtorum occurs on the Pacific slopes
of the Sierra Madre in Chiapas and Guatemala, where it occurs
with P. euthysanota euthysanota; P. schmidtorum chamulae is
known from only two localities on the Atlantic slopes of the
Central Highlands of Chiapas, where it occurs close to, but as
now known not with, P. euthysanota macrotympanum. On the
Atlantic slopes of the Sierra Madre Oriental in northern Oaxaca P.
ignicolor occurs with P. leonhardschultzei.
In the Sierra de los Tuxtlas in southern Veracruz and in the cloud
forests along the eastern slopes of the Sierra Madre Oriental northward
to Nuevo León, Hyla miotympanum seems to be the ecological
replacement of Ptychohyla. On the Pacific slopes north of
Guerrero, México, humid forests in which there are cascading mountain
streams are absent; consequently, no Ptychohyla are known
from that region. In the mountains of El Salvador Ptychohyla
euthysanota euthysanota occurs sympatrically with another small
stream-breeding hylid, Hyla salvadorensis. To the south of Honduras
the highlands diminish into the lowlands of Nicaragua, where
habitat suitable for Ptychohyla apparently does not exist. In the
mountains of Costa Rica and Panamá, the habitats occupied by
Ptychohyla in northern Central America are filled by a variety of
[Pg 342]
stream-breeding Hyla, such as Hyla legleri, H. rivularis, H. rufioculis,
H. alleei, and H. uranochroa.
Although members of the genus Ptychohyla occur in the southern
part of the Mexican Highlands to the west of the Isthmus of
Tehuantepec, the greater distribution and differentiation in the
genus is in the Chiapan-Guatemalan Highlands. In this respect
Ptychohyla is a counterpart of Plectrohyla.
Frogs of the genus Ptychohyla are ecologically associated with
mountain streams at elevations between 650 and 2200 meters; in
the geographic region where these frogs occur the vegetation between
those elevations consists of cloud forest or pine-oak forest.
In some places the frogs have been found in a mixture of oak and
semi-deciduous scrub forest. At Vista Hermosa, Oaxaca, P. leonhardschultzei
and P. ignicolor were found in cloud forest, whereas
at Agua del Obispo, Guerrero, the former species was found in pine-oak
forest. Ptychohyla schmidtorum is known only from cloud
forest; P. euthysanota euthysanota and P. spinipollex generally are
found in cloud forest, but in some places they live in pine-oak forest.
Ptychohyla euthysanota macrotympanum has been found in pine-oak
forest and in a mixture of oak and semi-deciduous scrub forest.
With the possible exception of the members of the Ptychohyla
schmidtorum group, which has been found only in cloud forest,
it seems as though the type of vegetation is not the controlling
factor in the ecological distribution of these frogs.
Ptychohyla has been found only where there are clear, cascading
streams overhung by vegetation, on which adults and young perch
at night, or even by day. The presence of these streams, in which
the tadpoles live, seems to be an important factor in the distribution
of Ptychohyla. As has been shown previously, the tadpoles of
Ptychohyla are adapted for existence in torrential streams, where
the water is cool, and the amount of oxygen is high. Clearly
these tadpoles are unsuited for life in ponds or sluggish streams in
the lowlands, where the temperature of the water is high, a layer
of silt on the bottom is deep, and the amount of oxygen is low. The
tadpoles cling to rocks on the bottom of the streams; there they
move slowly across the rocks, apparently feeding on the thin covering
of algae. Tadpoles were not observed on rocks having a thick
covering of algae or moss. The tadpoles were observed to swim
against the current in torrential streams, in which no fishes were
[Pg 343]
found. Therefore, it seems as though the presence of the stream-habitat
for the tadpoles is a significant factor in the ecological distribution
of the species of Ptychohyla.
At localities where two species of Ptychohyla occur sympatrically
(P. ignicolor and P. leonhardschultzei at Vista Hermosa, Oaxaca,
and P. euthysanota euthysanota and P. schmidtorum schmidtorum
at Finca La Paz, Depto. San Marcos, Guatemala) effort was made
to determine what, if any, ecological interspecific relationships
existed. Although adults of the sympatric species were found on
adjacent leaves or branches of bushes overhanging the streams at
both localities, segregation at the time of breeding seems to be
maintained by the notably different breeding calls in sympatric
species (see discussion of breeding calls). Thus, as has been shown
by Blair (1956), Fouquette (1960), and others working on a variety
of pond-breeding frogs and toads, the breeding call in Ptychohyla
acts as an important reproductive isolating mechanism.
At no locality were Ptychohyla and associated species of hylids
found so abundantly as were species of pond-breeding hylids in the
lowlands. Apparently reproductive activity is not concentrated in a
short breeding season, and it is highly doubtful if the populations
of these frogs are as large as those of the lowland pond-breeders.
The continual humid conditions and abundance of insect food
throughout the year in the cloud forest are perhaps indicative of
little interspecific competition among adults of Ptychohyla and
other sympatric hylids.
At Finca La Paz, Guatemala, tadpoles of two species of Ptychohyla
were ecologically segregated. The tadpoles of P. euthysanota
euthysanota were found in riffles in the streams, whereas those of
P. schmidtorum schmidtorum were found in slower water, chiefly
in small pools in the streams. At Vista Hermosa, Oaxaca, tadpoles
of P. leonhardschultzei were found in riffles, and tadpoles of the
sympatric P. ignicolor were found in a small pool in a stream.
Similar ecological relationships were observed for several species of
Costa Rican hylids. Throughout the range of Ptychohyla east of
the Isthmus of Tehuantepec, members of the genus occur with
species of Plectrohyla, all of which are larger than Ptychohyla, and
all of which have tadpoles that live in torrential streams. Tadpoles
of Ptychohyla spinipollex have been found in streams inhabited by
the tadpoles of Plectrohyla guatemalensis and P. quecchi; tadpoles
of Ptychohyla euthysanota euthysanota and P. schmidtorum schmidtorum
[Pg 344]
were found in streams inhabited by tadpoles of Plectrohyla
guatemalensis, P. matudai, and P. sagorum. In some streams great
numbers of tadpoles occur. The habitat is rather restricted, and
the food supply is limited. Consequently, interspecific competition
among the various species of hylids whose tadpoles live in the torrential
streams probably is highest during the larval stage. Unfortunately,
this aspect of salientian population ecology has received
no intensive study.
Since the cloud forests inhabited by Ptychohyla are daily bathed
in clouds and have a fairly evenly distributed rainfall throughout
the year, the frogs living in these forests are active throughout the
year. At least some of the species evidently have a long breeding
season, for I found calling males of P. leonhardschultzei in February,
March, and August, and found tadpoles in February, March,
June, and August. Tadpoles of the various species have been obtained
throughout much of the year, as follows: P. euthysanota
euthysanota, February, March, May, and July; P. euthysanota
macrotympanum, March, June, and August; P. spinipollex, February,
March, April, June, July, and August; P. schmidtorum
schmidtorum, March, May, June, July, and August; P. schmidtorum
chamulae, June and August; P. ignicolor, June. I suspect that this
temporal distribution more accurately reflects the seasonal activities
of collectors than of the frogs.
Calling frogs usually are on vegetation adjacent to or overhanging
streams; some calling males of P. spinipollex were on rocks in
or by streams. Clasping pairs of P. euthysanota and P. schmidtorum
were observed on vegetation by streams. Despite intensive
search, no eggs were found. It is doubtful if Ptychohyla deposit
eggs on vegetation overhanging streams, as do centrolenids and
Phyllomedusa, for egg-clutches of these frogs are easily found.
Possibly the eggs are laid separately on vegetation above the stream,
in which case they could be overlooked easily. In streams where
Ptychohyla and other hylid tadpoles occur, empty egg capsules
have been found on the lee sides of rocks, but there is no way to
determine which species laid the eggs.
Numbers of eggs were counted in gravid females; the largest eggs
have diameters ranging from 2.5 to 3.0 mm. The smaller species,
comprising the Ptychohyla schmidtorum group, have fewer eggs
than do the larger species. Numbers of eggs found in females of
the various species are: P. euthysanota euthysanota, 108; P. euthysanota
[Pg 345]
macrotympanum, 136, 160; P. leonhardschultzei, 141; P.
spinipollex, 119, 134, 143; P. schmidtorum schmidtorum, 59, 61, 90;
P. schmidtorum chamulae, 60, 71, 89.
Duration of the larval stage is unknown. Metamorphosing young
have been found from May through August. From two to six completely
metamorphosed young are available for each of the species,
except for P. ignicolor of which none is available. The smallest
young frog is a P. euthysanota having a snout-vent length of
14.2 mm.; the largest young frog is a P. schmidtorum schmidtorum
having a snout-vent length of 17.0 mm.
The preceding data on morphology, life histories, and behavior
form the basis for the following interpretation of the phylogeny of
Ptychohyla. Additional data are needed to support some of the
ideas discussed below; many of the data that are available for
Ptychohyla are lacking for other, possibly related, hylids. The
family Hylidae is composed of several hundred species, and most
of the species are poorly known. Consequently, any attempt to
place Ptychohyla in the over-all scheme of hylid phylogeny would
be premature at this time. But, as between the five species of two
species-groups here recognized as constituting the genus Ptychohyla,
some estimate of relationships can be made. First, it is necessary
to determine the validity of the genus itself.
As stated in the diagnosis of the genus, the only character that
sets this group of species apart from other hylids is the presence of
ventrolateral glands in the breeding males. To many systematists
the thought of being able to identify to genus only breeding males
is sufficiently disturbing to cause them to view with disfavor the
recognition of the genus. Nonetheless, the question is raised: Do
the five species herein placed in the genus Ptychohyla constitute a
natural assemblage? If the genus is considered to be more than a
category of convenience, that is to say, a group of related species
having a common origin, the primary problem is to determine
whether or not the five species form a phylogenetic unit.
The species of Ptychohyla are divided into two groups on the
basis of external morphology, breeding calls, and tadpoles. The
Ptychohyla euthysanota group seems to be a natural group composed
of three species, all of which are more closely related to one
another than to any other hylid. Likewise, the species comprising
[Pg 346]
the Ptychohyla schmidtorum group seem to represent a natural unit.
If the presence of ventrolateral glands in breeding males is ignored,
a student of salientian systematics might derive the Ptychohyla
euthysanota group from a hylid stock containing Hyla miotympanum
and Hyla mixomaculata. Likewise, Ptychohyla schmidtorum
could be placed with Hyla uranochroa and related species in
Costa Rica. Nonetheless, the fact remains that all of the species
assigned to the genus Ptychohyla have ventrolateral glands in the
breeding males; furthermore, ventrolateral glands are unknown in
other hylids. If the P. schmidtorum group and the P. euthysanota
group each evolved from separate hylid stocks, then the ventrolateral
glands must have developed independently in both groups.
That ventrolateral glands developed independently in five species
of frogs in southern México and northern Central America and not
in any of the other approximately 500 species of hylids in the world
is untenable. It is more logical to assume that the development of
the glands took place only once in a stock of hylids that gave rise
to the five species herein recognized as members of the genus
Ptychohyla.
The affinities of Ptychohyla apparently are not with any of the
other groups that have been generically separated from Hyla. Of
the daughter genera in Middle America only Plectrohyla has stream-adapted
tadpoles, but these large frogs are not closely related to
Ptychohyla. Stuart (1954:169) suggested that certain montane
species of Hyla in lower Central America and Hyla salvadorensis
in El Salvador may be related to Ptychohyla or even congeneric.
I have had experience with most of these species in the field and
believe that Stuart was correct in his suggestion of relationships.
The species concerned are four red-eyed stream-breeding Hyla in
Costa Rica—H. alleei, H. legleri, H. rufioculis, and H. uranochroa,
plus Hyla salvadorensis in the mountains of El Salvador. Morphologically
all of the species are similar; Hyla uranochroa, H.
legleri, and H. rufioculis have a lateral white stripe that is expanded
to form a spot beneath the eye, as in Ptychohyla schmidtorum. The
tadpoles of Hyla rufioculis and H. uranochroa have large funnel-shaped
mouths and long slender tails like those of Ptychohyla
schmidtorum. Lips of the tadpoles of H. legleri and H. salvadorensis
are folded laterally, in this respect resembling those of the Ptychohyla
euthysanota group. I do not know the tadpoles and the
breeding call of Hyla alleei. The breeding calls of Hyla rufioculis
[Pg 347]
and H. uranochroa consist of high melodious notes; the calls of
H. legleri and H. salvadorensis consist of series of short notes that
have the general characteristics of the call of Ptychohyla schmidtorum.
Affinities of the genus Ptychohyla seem to me to be with
the red-eyed species forming the Hyla uranochroa group in Costa
Rica. All of the species in the Hyla uranochroa group have large
frontoparietal fontanelles, rather small ethmoids, and small nasals
that are not in contact with one another or with the ethmoid. Some
species have a complete quadratojugal-maxillary arch; others do
not. Assuming that the parental stock that gave rise both to the
Hyla uranochroa group and to Ptychohyla was widespread in
Central America at a time of cooler, more humid conditions, it is
possible that with subsequent warming temperatures and seasonal
rainfall in the lowlands the parental stock was restricted to the
Costa Rican highlands, where the Hyla uranochroa group developed,
and to the Chiapas-Guatemala highlands, where Ptychohyla
evolved.
Ptychohyla schmidtorum is thought to resemble more closely the
parental stock of the genus than does any other species of Ptychohyla
now extant. This parental stock is discussed above in the
account of the generic relationships. Ptychohyla schmidtorum
has a red eye, white lateral stripe, frontoparietal fontanelle, funnel-shaped
mouth in tadpoles, and lacks nuptial spines; in all of these
characters it resembles members of the Hyla uranochroa group.
Probably during times of glaciation during the Pleistocene, when
climates in México and Central America were depressed, the
Ptychohyla stock was more widespread than it is now. Subsequent
elevation of climatic zones during interglacial periods would have
isolated populations as they are today in regions of cloud forests.
Thus, through geographic isolation populations could have differentiated
and evolved into the present species. Climatic fluctuation
in the Pleistocene must have been of sufficient magnitude to
permit the spread of cool, moist forests containing Ptychohyla across
the Isthmus of Tehuantepec into the mountains of Oaxaca.
Because of its small nuptial spines, small triangular vomers,
coloration, and absence of a rostral keel, Ptychohyla euthysanota,
more than any of the other species in the P. euthysanota group, resembles
P. schmidtorum. At the present time P. euthysanota and
P. schmidtorum are sympatric.
As I have mentioned previously, ecological segregation and
[Pg 348]
interspecific competition probably is highly developed in the tadpoles
of Ptychohyla. If this ecological segregation resulted from intraspecific
competition in a stock of Ptychohyla, possibly P. euthysanota
and P. schmidtorum differentiated sympatrically in this way.
Specific identity is maintained, at least in part, by different breeding
calls in males.
Ptychohyla spinipollex and P. leonhardschultzei seem to be more
closely related to one another than either is to P. euthysanota.
Probably a stock of P. euthysanota was isolated on the Atlantic
slopes of northern Central America from P. euthysanota on the
southern slopes. The frogs on the Atlantic slopes differentiated and
spread into the mountains of Oaxaca, where through isolation by
the barrier of the Isthmus of Tehuantepec they developed into P.
leonhardschultzei, while the stock on the northern slopes of Central
America evolved into P. spinipollex. Subsequent to the differentiation
of P. leonhardschultzei and P. spinipollex from P. euthysanota
and during a time of cooler more equable climate than exists now,
P. euthysanota and P. schmidtorum invaded the Central Highlands
of Chiapas. Subsequent climatic changes isolated populations of
each in the Central Highlands, where P. euthysanota macrotympanum
and P. schmidtorum chamulae evolved. Ptychohyla
ignicolor probably represents stock of P. schmidtorum that crossed
the Isthmus of Tehuantepec and became isolated in Oaxaca on the
western side of the isthmus.
[Pl 12]

Ptychohyla euthysanota euthysanota (KU 58008).
× 2.

Ptychohyla euthysanota macrotympanum (KU 58049). × 2.

Ptychohyla leonhardschultzei (KU 64117). × 2.

Ptychohyla spinipollex (KU 58054). × 2.

Ptychohyla schmidtorum schmidtorum
(KU 58043). × 2.
[Pl 17]

Ptychohyla schmidtorum chamulae (KU
58069). × 2.
[Pl 18]

Ptychohyla ignicolor (UMMZ 119603).
× 2.
1934. Über eine sammlung von Reptilien und Amphibien aus Mexiko,
Zool. Anz., 106:184-186, April 15.
1956. Call difference as an isolation mechanism in southwestern toads
(genus Bufo). Texas Jour. Sci., 8:87-106, March.
1956. The frogs of the hylid genus Phrynohyas Fitzinger, 1843. Misc.
Publ. Mus. Zool. Univ. Michigan, 96:1-47, pls. 1-6, February 21.
1960. Synonymy, variation, and distribution of Ptychohyla leonhard-schultzei
Ahl. Studies of American Hylid Frogs, IV. Herpetologica,
16:191-197, September 23.
1961. Descriptions of two species of frogs, genus Ptychohyla. Studies of
American Hylid Frogs, V. Univ. Kansas Publ. Mus. Nat. Hist.,
13:349-357, pl. 25, April 27.
1960. Isolating mechanisms in three sympatric treefrogs in the Canal
Zone. Evolution, 14:484-497, December.
1928. An apparently new Hyla from El Salvador. Proc. Biol. Soc. Washington,
41:123-124, June 29.
1952. Die Amphibien und Reptilien von El Salvador. Senckenbergischen
Naturf. Gesell., 487:1-120, pls. 1-16, December 1.
1912. Color standards and color nomenclature. Washington, D. C., 44
pp., 53 pls.
1951. Notes on a herpetological collection from Oaxaca and other localities
in Mexico. Proc. U. S. Nat. Mus., 101:465-484, May 17.
1954. Descriptions of some new amphibians and reptiles from Guatemala.
Proc. Biol. Soc. Washington, 67:159-178, August 5.
1957. Notes on a collection of amphibians and reptiles from southern
Mexico, with a description of a new Hyla. Great Basin Nat.,
17:52-56, July 31.
1937. New species of hylid frogs from Mexico with comments on the rare
Hyla bistincta Cope. Proc. Biol. Soc. Washington, 50:43-54, pls.
2-3, April 21.
1942. New tailless amphibia from Mexico. Univ. Kansas Sci. Bull., 28:
67-89, May 15.
1944. A new genus and species of Mexican frogs. Univ. Kansas Sci.
Bull., 30:41-45, June 12.
1949. New or unusual Mexican amphibians. Amer. Mus. Novitates,
1437:1-21, December 7.
Transmitted December 27, 1962.
[Pub_1]
MUSEUM OF NATURAL HISTORY
Institutional libraries interested in publications exchange may obtain this
series by addressing the Exchange Librarian, University of Kansas Library,
Lawrence, Kansas. Copies for individuals, persons working in a particular
field of study, may be obtained by addressing instead the Museum of Natural
History, University of Kansas, Lawrence, Kansas. There is no provision for
sale of this series by the University Library, which meets institutional requests,
or by the Museum of Natural History, which meets the requests of individuals.
Nevertheless, when individuals request copies from the Museum, 25 cents should
be included, for each separate number that is 100 pages or more in length, for
the purpose of defraying the costs of wrapping and mailing.
* An asterisk designates those numbers of which the Museum's supply (not the Library's
supply) is exhausted. Numbers published to date, in this series, are as follows:
| Vol. 1. | Nos. 1-26 and index. Pp. 1-638, 1946-1950. | ||
| *Vol. 2. | (Complete) Mammals of Washington. By Walter W. Dalquest. Pp. 1-444, 140 figures in text. April 9, 1948. | ||
| Vol. 3. | *1. | The avifauna of Micronesia, its origin, evolution, and distribution. By Rollin H. Baker. Pp. 1-359, 16 figures in text. June 12, 1951. | |
| *2. | A quantitative study of the nocturnal migration of birds. By George H. Lowery, Jr. Pp. 361-472, 47 figures in text. June 29, 1951. | ||
| 3. | Phylogeny of the waxwings and allied birds. By M. Dale Arvey. Pp. 473-530, 49 figures in text, 13 tables. October 10, 1951. | ||
| *4. | Birds from the state of Veracruz, Mexico. By George H. Lowery, Jr., and Walter W. Dalquest. Pp. 531-649, 7 figures in text, 2 tables. October 10, 1951. | ||
| Index. Pp. 651-681. | |||
| *Vol. 4. | (Complete) American weasels. By E. Raymond Hall. Pp. 1-466, 41 plates, 31 figures in text. December 27, 1951. | ||
| Vol. 5. | Nos. 1-37 and index. Pp. 1-676, 1951-1953. | ||
| *Vol. 6. | (Complete) Mammals of Utah, taxonomy and distribution. By Stephen D. Durrant. Pp. 1-549, 91 figures in text, 30 tables. August 10, 1952. | ||
| Vol. 7. | Nos. 1-15 and index. Pp. 1-651, 1952-1955. | ||
| Vol. 8. | Nos. 1-10 and index. Pp. 1-675, 1954-1956. | ||
| Vol. 9. | *1. | Speciation of the wandering shrew. By James S. Findley. Pp. 1-68, 18 figures in text. December 10, 1955. | |
| 2. | Additional records and extension of ranges of mammals from Utah. By Stephen D. Durrant, M. Raymond Lee, and Richard M. Hansen. Pp. 69-80. December 10, 1955. | ||
| 3. | A new long-eared myotis (Myotis evotis) from northeastern Mexico. By Rollin H. Baker and Howard J. Stains. Pp. 81-84. December 10, 1955. | ||
| 4. | Subspeciation in the meadow mouse, Microtus pennsylvanicus, in Wyoming. By Sydney Anderson. Pp. 85-104, 2 figures in text. May 10, 1956. | ||
| 5. | The condylarth genus Ellipsodon. By Robert W. Wilson. Pp. 105-116, 6 figures in text. May 19, 1956. | ||
| 6. | Additional remains of the multituberculate genus Eucosmodon. By Robert W. Wilson. Pp. 117-123, 10 figures in text. May 19, 1956. | ||
| 7. | Mammals of Coahulia, Mexico. By Rollin H. Baker. Pp. 125-335, 75 figures in text. June 15, 1956. | ||
| 8. | Comments on the taxonomic status of Apodemus peninsulae, with description of a new subspecies from North China. By J. Knox Jones, Jr. Pp. 337-346, 1 figure in text, 1 table. August 15, 1956. | ||
| 9. | Extensions of known ranges of Mexican bats. By Sydney Anderson. Pp. 347-351. August 15, 1956. | ||
| 10. | A new bat (Genus Leptonycteris) from Coahulia. By Howard J. Stains. Pp. 353-356. January 21, 1957. | ||
| 11. | A new species of pocket gopher (Genus Pappogeomys) from Jalisco, Mexico. By Robert J. Russell. Pp. 357-361. January 21, 1957. | ||
| 12. | Geographic variation in the pocket gopher, Thomomys bottae, in Colorado. By Phillip M. Youngman. Pp. 363-387, 7 figures in text. February 21, 1958. | ||
| 13. | New bog lemming (genus Synaptomys) from Nebraska. By J. Knox Jones, Jr. Pp. 385-388. May 12, 1958. | ||
| 14. | Pleistocene bats from San Josecito Cave, Nuevo León, México. By J. Knox Jones, Jr. Pp. 389-396. December 19, 1958. | ||
| 15. | New subspecies of the rodent Baiomys from Central America. By Robert L. Packard. Pp. 397-404. December 19, 1958. | ||
| 16. | Mammals of the Grand Mesa, Colorado. By Sydney Anderson. Pp. 405-414, 1 figure in text, May 20, 1959. | ||
| 17. | Distribution, variation, and relationships of the montane vole, Microtus montanus. By Sydney Anderson. Pp. 415-511, 12 figures in text, 2 tables. August 1, 1959. | ||
| [Pub_2] | 18. | Conspecificity of two pocket mice, Perognathus goldmani and P. artus. By E. Raymond Hall and Marilyn Bailey Ogilvie. Pp. 513-518, 1 map. January 14 1960. | |
| 19. | Records of harvest mice, Reithrodontomys, from Central America, with description of a new subspecies from Nicaragua. By Sydney Anderson and J. Knox Jones, Jr. Pp. 519-529. January 14, 1960. | ||
| 20. | Small carnivores from San Josecito Cave (Pleistocene), Nuevo León, México. By E. Raymond Hall. Pp. 531-538, 1 figure in text. January 14, 1960. | ||
| 21. | Pleistocene pocket gophers from San Josecito Cave, Nuevo León, México. By Robert J. Russell. Pp. 539-548, 1 figure in text. January 14, 1960. | ||
| 22. | Review of the insectivores of Korea. By J. Knox Jones, Jr., and David H. Johnson. Pp. 549-578. February 23, 1960. | ||
| 23. | Speciation and evolution of the pygmy mice, genus Baimoys. By Robert L. Packard. Pp. 579-670, 4 plates, 12 figures in text. June 16, 1960. | ||
| Index. Pp. 671-690 | |||
| Vol. 10. | 1. | Studies of birds killed in nocturnal migration. By Harrison B. Tordoff and Robert M. Mengel. Pp. 1-44, 6 figures in text, 2 tables. September 12, 1956. | |
| 2. | Comparative breeding behavior of Ammospiza caudacuta and A. maritima. By Glen E. Woolfenden. Pp. 45-75, 6 plates, 1 figure. December 20, 1956. | ||
| 3. | The forest habitat of the University of Kansas Natural History Reservation. By Henry S. Fitch and Ronald R. McGregor. Pp. 77-127, 2 plates, 7 figures in text, 4 tables. December 31, 1956. | ||
| 4. | Aspects of reproduction and development in the prairie vole (Microtus ochrogaster). By Henry S. Fitch. Pp. 129-161, 8 figures in text, 4 tables. December 19, 1957. | ||
| 5. | Birds found on the Arctic slope of northern Alaska. By James W. Bee. Pp. 163-211, plates 9-10, 1 figure in text. March 12, 1958. | ||
| *6. | The wood rats of Colorado: distribution and ecology. By Robert B. Finley, Jr. Pp. 213-552, 34 plates, 8 figures in text, 35 tables. November 7, 1958. | ||
| 7. | Home ranges and movements of the eastern cottontail in Kansas. By Donald W. Janes. Pp. 553-572, 4 plates, 3 figures in text. May 4, 1959. | ||
| 8. | Natural history of the salamander, Aneides hardyi. By Richard F. Johnston and Gerhard A. Schad. Pp. 573-585. October 8, 1959. | ||
| 9. | A new subspecies of lizard, Cnemidophorus sacki, from Michoacán, México. By William E. Duellman. Pp. 587-598, 2 figures in text. May 2, 1960. | ||
| 10. | A taxonomic study of the middle American snake, Pituophis deppei. By William E. Duellman. Pp. 599-610, 1 plate, 1 figure in text. May 2, 1960. | ||
| Index. Pp. 611-626. | |||
| Vol. 11. | Nos. 1-10 and index. Pp. 1-703, 1958-1960. | ||
| Vol. 12. | 1. | Functional morphology of three bats: Sumops, Myotis, Macrotus. By Terry A. Vaughan. Pp. 1-153, 4 plates, 24 figures in text. July 8, 1959. | |
| *2. | The ancestry of modern Amphibia: a review of the evidence. By Theodore H. Eaton, Jr. Pp. 155-180, 10 figures in text. July 10, 1959. | ||
| 3. | The baculum in microtine rodents. By Sydney Anderson. Pp. 181-216, 49 figures in text. February 19, 1960. | ||
| *4. | A new order of fishlike Amphibia from the Pennsylvanian of Kansas. By Theodore H. Eaton, Jr., and Peggy Lou Stewart. Pp. 217-240, 12 figures in text. May 2, 1960. | ||
| 5. | Natural history of the bell vireo. By Jon C. Barlow. Pp. 241-296, 6 figures in text. March 7, 1962. | ||
| 6. | Two new pelycosaurs from the lower Permian of Oklahoma. By Richard C. Fox. Pp. 297-307, 6 figures in text. May 21, 1962. | ||
| 7. | Vertebrates from the barrier island of Tamaulipas, México. By Robert K. Selander, Richard F. Johnston, B. J. Wilks, and Gerald G. Raun. Pp. 309-345, pls. 5-8. June 18, 1962. | ||
| 8. | Teeth of Edestid sharks. By Theodore H. Eaton, Jr. Pp. 347-362, 10 figures in text. October 1, 1962. | ||
| More numbers will appear in volume 12. | |||
| Vol. 13. | 1. | Five natural hybrid combinations in minnows (Cyprinidae). By Frank B. Cross and W. L. Minckley. Pp. 1-18. June 1, 1960. | |
| 2. | A distributional study of the amphibians of the Isthmus of Tehuantepec, México. By William E. Duellman. Pp. 19-72, pls. 1-8, 3 figures in text. August 16, 1960. | ||
| 3. | A new subspecies of the slider turtle (Pseudemys scripta) from Coahulia, México. By John M. Legler. Pp. 73-84, pls. 9-12, 3 figures in text. August 16, 1960. | ||
| 4. | Autecology of the copperhead. By Henry S. Fitch. Pp. 85-288, pls. 13-20, 26 figures in text. November 30, 1960. | ||
| 5. | Occurrence of the garter snake, Thamnophis sirtalis, in the Great Plains and Rocky Mountains. By Henry S. Fitch and T. Paul Maslin. Pp. 289-308, 4 figures in text. February 10, 1961. | ||
| 6. | Fishes of the Wakarusa river in Kansas. By James E. Deacon and Artie L. Metcalf. Pp. 309-322, 1 figure in text. February 10, 1961. | ||
| 7. | Geographic variation in the North American cyprinid fish, Hybopsis gracilis. By Leonard J. Olund and Frank B. Cross. Pp. 323-348, pls. 21-24, 2 figures in text. February 10, 1961. | ||
| [Pub_3] | 8. | Descriptions of two species of frogs, genus Ptychohyla; studies of American hylid frogs, V. By William E. Duellman. Pp. 349-357, pl. 25, 2 figures in text. April 27, 1961. | |
| 9. | Fish populations, following a drought, in the Neosho and Marais des Cygnes rivers of Kansas. By James Everett Deacon. Pp. 359-427, pls. 26-30, 3 figs. August 11, 1961. | ||
| 10. | Recent soft-shelled turtles of North America (family Trionychidae). By Robert G. Webb. Pp. 429-611, pls. 31-54, 24 figures in text, February 16, 1962. | ||
| Index. Pp. 613-624. | |||
| Vol. 14. | 1. | Neotropical bats from western México. By Sydney Anderson. Pp. 1-8. October 24, 1960. | |
| 2. | Geographic variation in the harvest mouse. Reithrodontomys megalotis, on the central Great Plains and in adjacent regions. By J. Knox Jones, Jr., and B. Mursaloglu. Pp. 9-27, 1 figure in text. July 24, 1961. | ||
| 3. | Mammals of Mesa Verde National Park, Colorado. By Sydney Anderson. Pp. 29-67, pls. 1 and 2, 3 figures in text. July 24, 1961. | ||
| 4. | A new subspecies of the black myotis (bat) from eastern Mexico. By E. Raymond Hall and Ticul Alvarez. Pp. 69-72, 1 figure in text. December 29, 1961. | ||
| 5. | North American yellow bats, "Dasypterus," and a list of the named kinds of the genus Lasiurus Gray. By E. Raymond Hall and J. Knox Jones, Jr. Pp. 73-98, 4 figures in text. December 29, 1961. | ||
| 6. | Natural history of the brush mouse (Peromyscus boylii) in Kansas with description of a new subspecies. By Charles A. Long. Pp. 99-111, 1 figure in text. December 29, 1961. | ||
| 7. | Taxonomic status of some mice of the Peromyscus boylii group in eastern Mexico, with description of a new subspecies. By Ticul Alvarez. Pp. 113-120, 1 figure in text. December 29, 1961. | ||
| 8. | A new subspecies of ground squirrel (Spermophilus spilosoma) from Tamaulipas, Mexico. By Ticul Alvarez. Pp. 121-124. March 7, 1962. | ||
| 9. | Taxonomic status of the free-tailed bat, Tadarida yucatanica Miller. By J. Knox Jones, Jr., and Ticul Alvarez. Pp. 125-133, 1 figure in text. March 7, 1962. | ||
| 10. | A new doglike carnivore, genus Cynaretus, from the Clarendonian Pliocene, of Texas. By E. Raymond Hall and Walter W. Dalquest. Pp. 135-138, 2 figures in text. April 30, 1962. | ||
| 11. | A new subspecies of wood rat (Neotoma) from northeastern Mexico. By Ticul Alvarez. Pp. 139-143. April 30, 1962. | ||
| 12. | Noteworthy mammals from Sinaloa, Mexico. By J. Knox Jones, Jr., Ticul Alvarez, and M. Raymond Lee. Pp. 145-159, 1 figure in text. May 18, 1962. | ||
| 13. | A new bat (Myotis) from Mexico. By E. Raymond Hall. Pp. 161-164, 1 figure in text. May 21, 1962. | ||
| 14. | The mammals of Veracruz. By E. Raymond Hall and Walter W. Dalquest. Pp. 165-362, 2 figures. May 20, 1963. | ||
| 15. | The recent mammals of Tamaulipas, México. By Ticul Alvarez. Pp. 363-473, 5 figures in text. May 20, 1963. | ||
| More numbers will appear in volume 14. | |||
| Vol. 15. | 1. | The amphibians and reptiles of Michoacán, México. By William E. Duellman. Pp. 1-148, pls. 1-6, 11 figures in text. December 20, 1961. | |
| 2. | Some reptiles and amphibians from Korea. By Robert G. Webb, J. Knox Jones, Jr., and George W. Byers. Pp. 149-173. January 31, 1962. | ||
| 3. | A new species of frog (Genus Tomodactylus) from western México. By Robert G. Webb. Pp. 175-181, 1 figure in text. March 7, 1962. | ||
| 4. | Type specimens of amphibians and reptiles in the Museum of Natural History, the University of Kansas. By William E. Duellman and Barbara Berg. Pp. 183-204. October 26, 1962. | ||
| 5. | Amphibians and Reptiles of the Rainforests of Southern El Petén, Guatemala. By William E. Duellman. Pp. 205-249, pls. 7-10, 6 figures in text. October 4, 1963. | ||
| 6. | A revision of snakes of the genus Conophis (Family Colubridae, from Middle America). By John Wellman. Pp. 251-295, 9 figures in text. October 4, 1963. | ||
| 7. | A review of the Middle American tree frogs of the genus Ptychohyla. By William E. Duellman. Pp. 297-349, pls. 11-18, 7 figures in text. October 18, 1963. | ||
| More numbers will appear in volume 15. | |||
This file was derived from scanned images. With the exception of the
list of typographical errors that were corrected below, the original
text is presented.
In the copy of the original, the Plates were grouped together between
pages 328 and 329. Here a smaller version of the illustration was placed
beside the text of the Systematic Account listing with a link to the
'full-sized' Plates which were moved to the end of the article's text
just above the listing of the Literature Cited. The Plate text contains
the notation "× 2" after the caption to let the reader know that the
image was enlarged by a factor of two. The images displayed will most
likely NOT be in 1:1 scale with the original printed Plates.
Typographical Errors Corrected:
Several minor typographical corrections were made (missing periods,
commas, misspelling of 'and', etc.); but are not indicated here. More
substantial changes are listed below:
References to Plate 11 (Audiograms): Pl. 1A, Pl. 1B, ...=> Pl. 11A, Pl. 11B, ...
References to the other Plates: Pl. 2, Pl. 3,... => Pl. 12, Pl. 13,...
Page 301, Paragraph 1: know => known
Page 302, Paragraph 1: Zoolegy => Zoology
Page 303, Paragraph 5: speces => species
Page 305, Paragraph 1: excresences => excrescences
Page 308, Paragraph 6: xiphisterum => xiphisternum
Page 316, Paragraph 3: with => width
Page 327, Paragraph 1: leonhard-schultzei => leonhardschultzei
to match remaining report text
Page 331, Paragraph 1: skelton => skeleton
Publication List Vol. 13, No. 8.: Decriptions => Descriptions
Featured Books

Notes on Philippine Birds Collected by Governor W. Cameron Forbes
The Pycnogonida.W. H. DALL. XIV.[14] The Mollusks.C. R. EASTMAN. VII.[7] The Sharks' Teeth.S. GARMA...
Read More
Geographic Variation in the Pocket Gopher, Thomomys bottae, in Colorado
arily in the Piñon-juniper,Ponderosa Pine, and Short Grass zones of Daubenmire (1943) but in somelo...
Read More
The Dog Crusoe and his Master
hort, is very harrowing, and, as it is intimately connected with Crusoe’s subsequent history, we w...
Read More
Mr. Dooley Says
hin he come back to Ar-rchey Road. No, sir, whin ourpeople grab hands at th' altar, they're hooked u...
Read More
Final Weapon
alikehad passed their day of usefulness.Not far from the ruined bridgesupport, Morely could see the...
Read More
The People of the Black Circle
claimed. 'They are no wiser thanthe leeches who are helpless! Nay, he dies and none can say why. He ...
Read More
Search the Sky
not since childhood. Ghost Townwas a wonderful place to play. “Tag,” “Follow My Fuehrer,”�...
Read More
Missing Link
The I-A section chief was dark-haired,gangling, with large headand big features. He stood in hiscust...
Read More
Comments on "A Review of the Middle American Tree Frogs of the Genus Ptychohyla" :